The easiest way to load a parameter file is by drag-and-drop from File Explorer, although you can certainly use the File: Open menu command, or the button on the main toolbar if you wish.
If you double-click a parameter file in File Explorer it will activate a new instance of Neurosim with the file pre-loaded.
Web Links
If you click on a web-link to a parameter file, you may be asked whether you want to download or run it. If you select run, be aware that a local copy of the file may be downloaded to your default Download folder, depending on your browser. To prevent disk clutter, you may want to delete these files after you finish your simulation session.
Each link activates a new instance of Neurosim, similar to double-clicking a local file.
In this Neurosim tutorial, parameter file links are shown in green thus.
Check the Measure box at the top of the Results view to open the Measure dialog.
If necessary, move thedialog so that it does not obscure the Results view.
The dialog is non-modal, so can be kept open while you do experiments
Drag the red cursor in the Results view to the X-location at which you want to make the measurement.
You can fine-position the cursor with the < and > buttons.
Click the Measure button in the Measure dialog. Note that the data value of each trace appears in the dialog in the labelled columns.
If there is more than one sweep, data are measured from each sweep in sequence unless a sweep is highlighted in the Results view, in which case only data from that sweep are measured.
Repeat the measurements at different X-locations as necessary.
Dual Cursors
You can measure values and their differences at two different times using dual cursors.
Check the 2 cursors box.
A blue cursor now appears in addition to the red cursor.
If you only want the difference between cursor values, check the Only difference box.
You can change which cursor is moved by the < and > buttons by selecting Red or Blue from the radio button choice.
User column
You can add a column to enter a user-specified value or label to each measurement row if desired:
Click the +User button to add a column at the left of the measurements.
Click the column header to edit its label if desired.
Click each cell in the user column and enter appropriate data.
You can navigate between rows with up- and down-arrow keys.
Plot
Click the Plot button to activate an X-Y scatter plot of the measurements.
Select which parameter goes on which axis from the axis drop-down list.
Context-sensitive help is available at any time within Neurosim by pressing F1.
Add horizontal and/or vertical cursors to the display by:
clicking the appropriate Results toolbar button ( or ), or
selecting from the View: Cursors menu, or
right-clicking in the Results view and selecting from the context menu.
Position cursors by dragging with the mouse.
You can adjust the fine position of a selected cursor using the keyboard arrow keys. A cursor is selected by clicking it. A selected cursor is indicated by the blue bar at its end.
The time (vertical cursor) or trace value (horizontal cursor) at the cursor location shows at its top or left end. The value labels can be dragged along the cursor to a more convenient screen location if desired. IMPORTANT: for horizontal cursors, the value is that for the axis range in which the cursor is located.
If you have multiple cursors displayed they can be coupled together so that when you move one cursors, all cursors of the same type move by the same amount. This option is available on the View: Cursors menu.
Cursors can be deleted by right-clicking within the Results view, and selecting the appropriate command from the context menu.
The transmembrane current carried by ions flowing through an ion channel is given by the following equation:
I = g(Vm-Veq)
where I is the current, g is the conductance of the channel to the ion, Vm is the membrane potential, and Veq is the equilibrium potential of the ion in question.
The factor (Vm - V eq) is known as the driving force, because the further the membrane potential is from the equilibrium potential of an ion, the greater the “push” on the ion resulting from the imbalance in the Nernst equation.
This equation is a simple variant of Ohm's Law (current = conductance x voltage), but voltage is represented by the driving force. It is one of the most important equations in cellular electrophysiology because it allows us to quantifiy current flow in both action potentials and chemical synapses.
Four different recording methods. a. Intracellular recording usually involves a microelectrode - a glass tube drawn out to a very fine point and filled with an electrolyte. The blunt end of the tube is connected by a wire to a recording amplifier or stimulator. b. A sharp microelectrode penetrates into the interior of the cell. This can record the membrane potential or inject current. c. A patch electrode attaches to the cell surface and breaks the membrane just under its tip. This can record and stimulate like method a, but usually causes less damage to the cell. Also a patch electrode is usually blunter than a sharp electrode, and passes current more easily and records with less noise. d. A patch electrode attaches to the cell surface with just a single ion channel under its tip. This can record the current through that channel, with the rest of the cell still attached and undamaged. e. As c, but the electrode is withdrawn from the cell, bringing the single channel with it. Current flowing through the channels can be recorded, and the chemical environment on both sides of the channel can be controlled by the experimenter. By careful manipulation the channel can be arranged to have its inner surface within the electrode lumen (outside-out patch) or its outer surface within the lumen (inside-out patch).
Many transfer functions of the form y = f(x) allow the user to choose between linear, sigmoid, and user-specified types. In each case there may be a specified minimum and maximum output.
Linear: There is linear interpolation between the minimum and maximum. Two parameters control the function:
Threshold: Below this input level, the output is the minimum.
Saturate: Above this input level, the output is the maximum.
For input between the threshold and saturate levels, the output scales linearly:
\begin{equation}
y = min + (max - min) \times (x - thresh) / (sat - thresh)
\label{eq:eqLinearTrF}
\end{equation}
Sigmoid: The function has a sigmoid shape varying between the minimum and maximum. Two parameters control the shape of the sigmoid:
Mid: The input value that produces output mid way up the curve of the sigmoid shape.
Slope: The steepness of the sigmoid curve. Often the inverse slope is used, so larger values produce shallower slopes.
\begin{equation}
y = min + (max - min)/(1+e^{\frac{mid-x}{slope}})
\label{eq:eqSigmoidTrF}
\end{equation}
User: The user specifies an equation using the built-in expression parser. Context-dependent parameters may be passed to the equation. It is up to the user to ensure that the output is appropriate for the intended use.
Select 0 (new) in the Experimental Contol panel.
The panel is normally docked on the left of the Setup view, but it can be floated, or docked elsewhere if desired.
Edit the stimulus parameters as desired.
Enter the appropriate ID number(s) into the Target neuron(s) box. You can apply the same stimulus to multiple neurons by entering multiple numbers.
Select an Existing Stimulus
Either Click on the appropriate square box attached to the target neuron in the Setup view
Or
Click on the stimulus number in the Stimulus list in the Experimental Control panel.
Edit an Existing Stimulus
Select the stimulus as above.
Edit the parameters in the Stimulus list in the Experimental Control panel.
Change the Target of an Existing Stimulus
Either Drag the square stimulus box from the existing target neuron to the new target neuron in the Setup view.
Or
Select the stimulus number in the Stimulus list in the Experimental Control panel.
Check the Measure box near the top of the Results view to activate the Measure dialog.
Move the dialog out of the way, so that you can see the Results.
In the Result view, set the Hilight sweep to 1. Now the largest spike is highlighted and the others are greyed out.
When a sweep is highlighted, measurements are only made from that sweep. If no sweep is highlighted, measurements are made from all sweeps simultaneously. We do not want the latter, because the location of the peak of the action potential varies in each sweep.
Click the expand timebase button in the Results toolbar (). This just makes it a bit easier to detect the peak of the spike. Drag the red vertical measurement cursor to line up with the peak of the spike.
Click the Measure button in the Measure dialog, and note that the membrane potential value at the spike peak appears in the Voltage column in the dialog.
Set the Hilight sweep to 2, move the vertical red cursor to the new highlighted spike peak, and repeat the measurement.
(Note: as well as dragging with the mouse, you can use the keyboard arrow keys to move the cursor, or click the <,> buttons in the dialog.)
Repeat for each sweep.
At this point you should have 5 rows of measurements in the dialog, with the spike peak at each concentration listed in the voltage column.
Click +User in the Measure dialog. This inserts a column at the left in which you can enter any data you like.
Click the left-most column header (the new user column) and enter "[Na]ext" in the Column header dialog. This is not strictly necessary, but it keeps things clearer.
Click in the first row of the user column, and enter 836.
Click in the second row (or press the down-arrow on the keyboard) and enter 418.
Repeat for the remaining rows, entering the appropriate concentration value.
Task: Plot the peak of the action potential against the log (base 10) of external the sodium concentration. You could just click Copy in the dialog, and paste the measurements into an external graphing program like Excel. However, we can get a quick preview of the graph in Neurosim itself:
Click the Plot button in the Measure dialog, to open the XY Scatter dialog.
In the new dialog, select Voltage from the Y axis dropdown list. You should now see a plot with data points curving upwards to the right.
Check the Log X box in the Plot dialog.
Check the Linear trendline box in the dialog, and note the value of the slope that appears under the check box.
The task is now completed.
Spikes are the mechanism that allow neurons to pass information over relatively long distances within themselves. Thus a mechanosensory neuron at the tip of a flipper in a blue whale has to send signals all the way to the spinal cord, and then up the cord to the brainstem. This is a long way, and passive conduction of the sort described previously could not do the job due to the attenuation of the signal with distance. Spikes are the answer!
Our understanding of how nerves generate spikes derives in large part from the seminal work of Hodgkin and Huxley. They won the Nobel prize for developing a model (the HH model) which is based on plausible biological mechanisms and which could be expressed mathematically with experimentally-derived parameters (Hodgkin and Huxley, 1952). Many of the simulations in Neurosim are based on this model.
The Hodgkin-Huxley model for generating the action potential builds on the RC model of the passive membrane given earlier, by including additional channels with variable conductances. Specifically, action potentials arise as a result of voltage- and time-dependent changes in the conductance of channels that are selectively permeable to sodium and potassium ions.
The membrane as an RC circuit. The circuit includes sodium, potassium and leakage ion channels.
The batteries represent the equilibrium potential for each set of ions.
A depolarising change in membrane potential from the resting condition causes two effects; a rapid but transient increase in the conductance of sodium channels, and a delayed increase in the conductance of potassium channels. The bigger the depolarisation, the bigger the increase in conductance (up to a maximum level).
The increase in sodium conductance in the resting neuron causes an inflow of sodium ions (a sodium current) according to the following variant of Ohm’s law:
where I is the current, g is the conductance to the ion (in this case sodium, but this is a general equation that can apply to any ion), Vm is the membrane potential, and Veq is the equilibrium potential of the ion. The factor (Vm - Veq) is known as the driving force, because the further the membrane potential is from the equilibrium potential of an ion, the greater the “push” on the ion resulting from the imbalance in the Nernst equation. Since the resting membrane potential is a long way below the sodium equilibrium potential,
there is initially a large inward driving force on sodium.
Sign convention
The sign convention for results displayed in Neurosim is that an inflow of positive ions is drawn as a negative current,
and the driving force equation \eqref{eq:eqDrivingForce} reflects this.
This is a bit counter-intuitive, but it fits with the standard way of displaying voltage clamp data in the literature.
Internally within the calculation engine, an inflow of positive ions of course causes depolarization.
If the initial increase in sodium conductance is big enough, it will become regenerative (self-increasing). This is because the inflow in positively-charged sodium ions (the inward current) itself leads to further depolarization, which causes a further increase in sodium conductance, and a further depolarisation of membrane potential etc. This is an example of positive feedback,
and it generates the rising phase of the action potential.
The sodium cycle. Voltage-dependent sodium channels mediate positive feedback.
However, the sodium conductance increase is intrinsically self-limiting in duration (i.e. it is transient), and so even though the membrane is very depolarised at the peak of the action potential, the sodium conductance starts to return to its resting level (a process known as inactivation) and the membrane starts to repolarize.
At about the same time that sodium conductance starts to inactivate, the delayed increase in potassium conductance leads to an outflow of potassium ions, which thus contributes to the falling phase of the action potential. This is negative feedback – the potassium outflow counteracts the depolarization that caused it in the first place. The increased potassium conductance shifts the membrane potential towards the potassium equilibrium potential, which is usually more negative than the resting potential, leading to the afterhyperpolarisation. Finally, the increase in potassium conductance is itself terminated as the depolarisation which initiated it is reversed, and the membrane returns to its original resting potential. At this point the sodium conductance has recovered from inactivation, and is ready to increase again if another depolarizing stimulus comes along.
The potassium cycle. Voltage-dependent potassium channels mediate negative feedback.
The HH model provides a predictive mathematical description of these events, and suggests a plausible set of biophysical mechanisms that might give rise to them.
Static (Non-Propagating) Spikes
Threshold: All or None
We will start with one of the simplest and most well-known properties of nerve impulses – they are famously “all or none”.
Load the parameter file Threshold from the Spikes sample sub-folder.
Click Start to run a simulation.
A very small stimulus is applied to the neuron (visible in the green lower trace), and it generates a tiny voltage response (the red upper trace).
Click the Run on change checkbox in the Results view. This is a convenience function that means that whenever a parameter changes, the simulation runs as if you had just clicked Start.
In the Setup view, click the up spin button of the Amp Pulse 1 parameter to increase the stimulus by a step of 5. (You could just manually edit the parameter value, but the spin button is easier.)
A slightly larger stimulus generates a slightly larger response, but essentially nothing much changes.
Click the spin button repeatedly, observing the voltage profile in the Results view. There should be a dramatic change when you reach a stimulus value of 30.
The stimulus has now crossed threshold and is generating a spike.
Click the spin button three more times.
The spike occurs slightly earlier with each larger stimulus, but does not change much in amplitude.
Click Clear.
Edit the Amp value to return it to 30 (the first value that generated a spike).
Right-click the Amp spin button to open the Spin Control Properties dialog.
Set the Delta value to 1. This controls the size of the change produced by a single click on the button.
Click OK to dismiss the dialog.
Now repeatedly click the down spin button to reduce the stimulus.
The spike collapses at a particular threshold stimulus.
Note that the extracellular sodium concentration shown in the Setup View is 418 mM, which is the concentration in sea water originally used by Hodgkin and Huxley.
Change the extracellular sodium concentrations to the following values, running an experiment at each value by clicking Start, without clearing the screen between runs:
You may also notice that the spike peak height decreases in approximately equal-sized steps,
although the changes in concentration are definitely not equal - they are halving at each step.
This immediately implies that there is some sort of logarithmic relationship between sodium concentration and peak spike amplitude.
Task: Plot the peak of the action potential against the log (base 10) of external the sodium concentration.
The first job is to measure the spike peaks. There are lots of ways to measure in Neurosim, but in this case the quickest and easiest is to use a horizontal cursor.
Right-click the Results view near the top of the biggest spike, and select Add horz cursor from the context menu.
[If necessary, you can delete a cursor by right-clicking it and choosing appropriately from the context menu.]
You can also add cursors by clicking buttons at the right-hand end of the Results toolbar (horizontal: , vertical: ), or with options in the View: Cursors menu.
Drag the cursor to line up precisely with the top of the biggest spike (when the concentration was 836 mM).
You can fine-tune the cursor position using the keyboard ↑ or ↓ keys, but you need to select it first by clicking on it.
For convenience, drag the measurement text above the cursor to the right, to get it clear of the spikes.
Plot a point on the graph below using the concentration at which the spike was generated to determined the X value and the cursor voltage read-out to determine the Y value.
You can right-click the graph, select Copy Image (the exact text may vary depending on your browser), and paste the image into a simple drawing program like Paint or PowerPoint. Then drawIn PowerPoint, the easiest way is to draw a small circle anywhere, and then drag it to the correct position on the graph. You can then copy, paste and drag the circle for subsequent points. a circle on the graph to mark the point.
Note that the graph is semi-logarithmic: the Y axis (voltage) has a standard linear scale, but the axis X (concentration) has a logarithmic scale. The vertical grid lines are thus drawn at the following values: 10 20 30 40 50 60 70 80 90 100 200 300 400 500 600 700 800 900 1000
You must use judgement to plot intermediate values, but with care you will not be far out.
Repeat for each spike.
The points should fall more-or-less on a straight line. Draw a trendline through the points (by eye is fine, no need to calculate a regression line).
Semi-log graph for plotting spike height against extracellular sodium concentration.
Question: For a 10-fold change in concentration (e.g. from 60 to 600 mM), what is the change in spike amplitude?
What can we learn from this graph? The most obvious thing is that the points fall on a (more-or-less) straight line. You will remember that the Nernst equation tells us that the equilibrium potential of an ion is directly proportional to the logarithm of the concentration gradient of that ion across the membrane, and that the constant of proportionality is 58For a monovalent ion, at room temperature, using log base 10, with the values in millivolts.. When we change the external sodium concentration we are changing that gradient for sodium, since the internal sodium concentration is not changed. You may also remember from the Goldman equation that the membrane potential approaches the equilibrium potential for an ion only when the membrane is much more permeable to that ion than any other.
[Note that the relationship is linear whether the logarithm is base 10 or base e (the natural logarithm), although of course the position of values on the X axis and hence the slope change between the two choices.]
Take-home message: When we plot the spike peak potential against the log of the exernal sodium concentration and get a straight line with a slope similar to the Nernstian proportionality constant, we are showing that at the time of the spike peak the membrane is far more permeable to sodium than to any other ion.
Congratulations, you have just done one of the classic experiments in early neurophysiology which demonstrated the sodium-dependence of action potentials.
Click the Clear button to clear the Results screen.
Set the external sodium concentration to 64 mM, which is the same as the intracellular concentration.
Click Start and measure the spike peak.
Question: Is the peak value 0? Why might you expect it to be? If it is not, how do you interpret that?
The only trace that is visible is the membrane potential, and absolutely nothing interesting happens because conditions are normal and there is no stimulus.
This simulation is based on the squid giant axon, where the normal extracellular potassium concentration is 10 mM (the concentration in seawater). However, the basic mechanism of spike generation is the same as in humans, as are many of the consequences of elevated potassium.
Click the up-arrow for the extracellular potassium to increase its concentration to 12 mM.
Repeatedly click the up-arrow until the concentration reaches 32 mM.
Question: Why does the neuron start to spike as you increase extracellular potassium concentration? Why does the neuron eventually stop spiking?
There are now 4 axes showing in the Results view. The top two show the standard membrane potential and stimulus respectively. The third shows the trans-membrane current carried by sodium and potassium, while the bottom axis shows the sodium and potassium conductance. Note that the current and conductance values cannot be measured directly in a real experiment of this kindCurrent can be measured in a voltage clamp experiment, and we will come to this later. – but one advantage of modelling is that it enables us to view these “behind the scenes” parameters.
Select the View:Trace ID: Show menu option.
A list of identifying abbreviations appears under the buttons on the left of the Results view, one for each trace. Make sure you understand what the abbreviations refer to; volt is the membrane potential, iStim, iNa, and iK, refer to the stimulus current, sodium current and potassium current respectively, gNa and gK refer to the sodium and potassium conductance.
Task: Use the mouse to drag these items to appropriate points on the screen to label the traces correctly. [The colour coding should help!]
Notice that the conductance traces are all positive – they start from a very low baseline, and increase and then decrease again. In contrast, the current traces are both positive and negative. By convention, an inward flow of positive ions is drawn as a negative current, while an outward flow of positive ions is drawn as a positive current. [This is somewhat confusing, but blame Hodgkin & Huxley, who got the Nobel prize for figuring this all out, so I’m not going to argue.] So which currents are inwards and which outwards during the action potential?
Examine the traces and see if you understand what is going on in the current and conductance traces in relation to the spike shown in the voltage trace. It may be useful to add a vertical cursor to help determine the relative timings of events
Questions: Here are some points to think about;
Which conductance increases first, the sodium or potassium?
Which conductance increase lasts longer, the sodium or potassium?
Question: With TEA applied, why is the sodium current so small, despite the large sodium conductance and the big voltage change? (Continue to think about the driving force.)
Molecular Events and Patch Recordings
Three key insights of Hodgkin and Huxley, which are now very well established but which were by no means obvious at the time, are
Sodium and potassium flow through separate populations of ion channels
The overall membrane conductance is the sum of these individual unitary conductances
The probability of an individual ion channel being open is voltage- and time-dependent.
They further proposed that each ion channel behaved as if it had a series of “gates” in it, and that for the channel to be open, all the gates within it had to be open simultaneously. Each gate flickered open and shut randomly, but the probability of it being open changed with the membrane potential. It was this voltage dependence of the gates that gave the channel as a whole its voltage dependence.
With this basic model structure, they found that they could produce a good fit to experimental data if they assumed that the sodium channel had 3 activation gates (called m-gates) which responded rapidly to voltage changes and which increased their open probability with depolarization, and one inactivation gate (the h-gate) which responded slightly more slowly, but decreased its open probability with depolarization. They also assumed the potassium channel had 4 activation gates (n gates), but these responded relatively slowly to voltage changes. Each gate was independent of the others, but all responded in a probabilistic way to voltage changes (see Advanced Kinetics and Implementation Details: Voltage dependent channels for a more detailed explanation).
The HH gate model. The gates are in a typical state for the resting potential.
This all makes sense at a qualitative level. Depolarization causes a rapid opening of m gates allowing an increase in sodium conductance and the depolarizing phase of the action potential, but this is soon followed by the closing of h gates causing the sodium conductance to fall again. This is sodium inactivation. The relatively slow activation of n gates in potassium channels cause a delayed increase in potassium conductance, which, along with sodium inactivation, terminates the action potential. The sodium h gates then re-open, but this does not happen immediate, so there is an absolute refractory period during which a second depolarization cannot open sodium channels. Furthermore, the potassium n gates are even slower to de-activate, causing an extended period of increased potassium conductance after the spike, which is the relatively refractory period.
Terminology
Neurophysiologists usually make the following distinctions:
Activation and de-activation are the opening and closing of activation gates, i.e. gates whose open probability increases with depolarization (e.g. m or n gates).
Inactivation and de-inactivation are the closing and opening of inactivation gates, i.e. gates whose open probability decreases with depolarization (e.g. h gates).
There is thus a difference between inactivation and deactivation.
Single Channel Patch Recording
Modern techniques allow recording current through single ion channels in a very small patch of membrane (a single-channelpatch recording), and these confirm the first three insights described above.
The top axis shows the membrane potential with an action potential, and the second shows the stimulus that initiates it. The third axis shows sodium and potassium conductance. The fourth axis shows the probability values of the individual gates. The red trace is the m gate (sodium activation), and this goes up (increased open probability) during the action potential. The green trace is the h gate (sodium inactivation), and this goes down during the action potential, but lags somewhat behind the m gate trace. The dark blue trace is the n gate (potassium activation), and this acts like the m gate, but again lags behind it.
In this simulation we have two patch electrodes in addition to the standard voltage recording. The fifth axis shows the conductance of a single sodium channel randomly selected from the whole population of sodium channels, and the sixth (bottom) axis shows the conductance of a single potassium channel, again randomly selected. (Note that conductance values are normalized so the conductance of a single open channel is 1.)
Channels open and close probabilistically, so I DON’T KNOW exactly what happened in the single channel recordings when you ran the experiment. The traces may well both be completely flat, indicating that in this particular run of the simulation, neither channel opened.
Repeatedly click Start and keep on eye on the bottom two traces. These are the only ones that might vary between runs. (Note Auto clear has been selected, so you don’t have to clear the screen between runs.)
You should start to see brief periods when the channels open (there is a square step upwards in the trace). These openings should be concentrated in the part of the sweep encompassing the action potential itself, in which the conductance traces (3rd axis) indicate that the conductance is relatively high.
The relatively low probability of channel opening during an individual action potential is at first quite surprising, but makes sense in terms of the model if you look at the numerical conductance values. The Maximum sodium conductance (visible in the Membrane Properties box in the Setup view) shows that the sodium conductance if all the sodium channels were open would be 120 mS. However, the peak value of the sodium conductance trace is about 33 mS, meaning that only about one third of the possible sodium channels are open at the peak of the action potential. A similar argument can be used for the potassium channels.
Micro and Macro Conductance
Increase the Patch channel Count in the Setup view to 5. This means that each patch electrode is now recording from a patch of membrane containing 5 randomly selected channels of its type.
Once again, click Start repeatedly.
Now it is likely that you will see some channel opening in most runs. You may well see several steps in the patch trace, indicating opening of multiple channels. You cannot specify which of the 5 possible channels opened for any particular step, you just know that an extra channel opened for each step.
Increase the Patch channel Count to 50 and click Start.
Hold down the control key and click the autoscale button in the Results toolbar to rescale the axes.
Now the patch recordings should start to look like the overall conductance traces in terms of their shape.
Take-home message: The overall sodium and potassium conductance expressed by the whole neuron is made up of large number of fixed-size single channel micro-conductances, which open and close probabilistically.
The HH Model in Action
An animated illustration of the HH model is available.
A new Membrane Channel Model window opens showing a visual representation of the HH channel model, with a sodium channel on the left and a potassium channel on the right.
Move the window so it does not obscure the Results.
Select a Slow down factor of 10 from the drop-down list in the main toolbar. This will enable you to see the cartoon animation more easily. You may need to adjust this value depending on the speed of your computer.
Click Start, and observe how the individual gates in the cartoon open and shut.
The gates open and close randomly, but with a probability that depends on the membrane potential at that time. Thus the m and n gates are more likely to be open during the peak of the action potential, while the h gate is more likely to be open at the resting or hyperpolarized potential.
When all the gates are open an arrow is drawn through the channel to show that it is now conducting. This will coincide with an open step in the patch traces in the Results.
Click Start repeatedly, until you have a good "feel" for how the gates open and close.
When you are ready, add TTX to the preparation, and click Start.
The sodium channel is blocked, so even if it opens, no sodium current can flow. The potassium channel is not blocked, but is unlikely to open because the membrane does not depolarize much.
Remove the TTX, add TEA, and click Start.
Now the voltage-dependent potassium channels are blocked and their conductance remains at 0 throughout. The depolarizing phase of the spike occurs, but then the sodium channels inactivate and the sodium conductance drops. However, the membrane potential does not return to the resting level. This is because the value of h stabilises at a low but non-zero level, and since the value of m is high, there is some residual sodium conductance (you could use the Measure facility to confirm this). This means that the final membrane potential is set by Goldman-type balance between the leakage conductance, and the low but non-zero conductance of the voltage-dependent sodium channels.
Remove the TEA and add Scorp Tx and click Start.
Scorp Tx stands for scorpion toxin. Amongst many other nasty components, the alpha-toxin in scorpion venom binds to the part of the sodium channel that mediates inactivation, and slows or prevents it. In the simulation the venom just locks the h gate open, which produces the same effect but is easily reversible!
The functional effect of scorpion toxin on the membrane potential is somewhat similar to that of TEA, but if you look at the model you will see that all the gates are wide open, and both types of channel are in a permanent high conduction state. It is not surprising that scorpion venom has such unpleasant effects on its victims.
Strength-Duration Curve
We know from personal experience that there is often some sort of trade-off between the strength of a stimulus and its duration – a very brief stimulus might be barely noticed, but if it lasts longer, it becomes more obvious (if it lasts for a very long time we might become used to it and stop noticing it altogether, but that’s another story).
This trade-off is well established down to the single neuron level, where it can be investigated by finding the stimulus strength that is just sufficient to elicit an action potential, for different durations of stimulus.
Start a graphing program like Excel or something similar.
Note that this uses the Advanced HH model, so the interface is slightly different from the standard HH model used previously.
A stimulus of strength 1 nA is applied for a duration of 20 ms. This does not elicit an action potential – the stimulus is below threshold.
Repeatedly click the up-arrow of the Pulse 1 stimulus Amplitude, until a spike occurs.
With each click, the stimulus strength increases by 0.1 nA. The run-on-change and auto-clear options have been selected, so a separate experiment runs with each click. A spike should occur when the strength reaches 1.7 nA (but note that there are “wobbles” in the membrane potential before that, indicating that some voltage-dependent processes are being activated even below threshold).
Enter the stimulus duration and threshold strength (20 and 1.7 respectively) into the spreadsheet.
Successively reduce the stimulus duration to 10, 5, 2, 1, 0.5, 0.2 and 0.1 ms.
At each duration:
Measure the threshold (the current strength which is just sufficient to elicit a spike). You should measure to 0.1 nA accuracy.
Enter the data into the spreadsheet.
Task: Plot a scatter graph of stimulus duration (X axis) against threshold strength (Y axis).
You should end up with a graph that has a general shape something like this:
Threshold Strength-Duration. There is a trade-off between the strength and duration of a just-threshold stimulus.
Take-home message: Long duration stimuli do not need to be as strong as short-duration stimuli to elicit an action potential, but there is a minimum strength below which action potentials cannot be elicited, no matter how long the stimulus.
Rheobase and Chronaxie
The minimum stimulus amplitude required to elicit an action potential when the stimulus has a long duration is called the rheobase of the neuron. A related value is the chronaxie, which is the minimum stimulus duration required to elicit an action potential when the stimulus amplitude is twice that of rheobase. Together, these numbers are helpful parameters that characterize the general excitability of a neuron. They are useful in many clinical studies, because various neurological diseases can alter their values due to underlying cellular pathologies.
Question: What are the rheobase and chronaxie values for the model neuron?
Cause of the Strength-Duration Relationship
One of the chief factors underlying the shape of the strength-duration curve is simply the RC properties of the passive membrane. We can investigate this by blocking the voltage-dependent channels.
Reload the file Strength Duration to return to the starting conditions.
Set the Pulse 1 stimulus Amplitude to 1.7, which we found to be just above threshold in the previous experiment, and which should generate a spike.
Click the TTX and TEA boxes in the Drugs frame of the Setup view. This blocks the voltage-dependent channels and prevents spiking, so the membrane becomes purely passive.
Note that when the simulation auto-runs as you apply the drugs, there is now an initial depolarization that precedes the onset of the stimulus. This is because some voltage-dependent channels are normally open even at resting potential, and blocking them causes a small depolarization. We can take account of this by changing the starting resting potential.
Click the Passive box in the Cell properties frame of the Setup view to open the Passive Properties dialog.
Set the Initial membrane potential to -60 mV. This is the equilibrium potential of the leakage channels, and this will be the stable membrane potential in the absence of voltage-dependent channels.
Click OK to dismiss the dialog. Note that the initial depolarization in the Results view has now disappeared – the only remaining response is due to the stimulus.
To increase the visibility of the data, set the top scale of the voltage axis to -40 and the bottom scale to -60 mV. You should now see a classic standard RC-type response.
We are going to assume that the maximum (final) voltage achieved within the 20 ms stimulus represents the voltage threshold of the neuron. Obviously, there is no actual threshold since there are no spikes, but it gives us a suitable datum point. The question is, what stimulus strength is needed to lift the membrane potential to this value, if we reduce the stimulus duration?
Click the Horizontal cursor button () in the Results toolbar.
Drag the cursor so that it lines up with the maximum voltage. You can use the keyboard up/down arrow keys for fine positioning. [From now on, do not change the size of the Results display, since that would change the cursor relative location.]
Enter the stimulus duration and threshold strength (20 and 1.7 respectively) into new columns in the spreadsheet.
Now reduce the stimulus duration to 2ms (a factor of 10 shorter).
Note that the voltage starts to rise at the same rate as before, but the stimulus switches off long before the response reaches the horizontal cursor. As soon as the stimulus switches off, the voltage starts to fall again. The only way to reach the cursor within the time available is if we can increase the rate of rise of the voltage.
Increase the Amplitude of the stimulus to 3 nA.
From the basic equation defining capacitance, an increase in current should result in an increase in the rate of change of voltage, and that is what we see: the voltage rises more steeply and reaches a higher level within the time of the stimulus.
Repeatedly click the up-arrow of the Pulse 1 stimulus Amplitude, until the peak voltage just reaches the horizontal cursor.
Enter the stimulus duration and amplitude into the spreadsheet.
Task: Find the threshold amplitude for a range of stimulus duration values to match those of the previous exercise involving spike threshold, and construct a strength-duration graph for the purely passive membrane. How does its shape compare to that of the spiking membrane?
Question: From your knowledge of RC properties, what factor determines the rheobase of the passive membrane?
Hopefully, this has given you insight into one of the main reasons why the spike threshold strength-duration curve has the shape that it has. However, in a spiking neuron there is more going on than just the RC properties - there are voltage-dependent channels that may open and/or inactivate during the sub-threshold part of a stimulus, and this can affect the shape of the strength-duration curve as well. This can be seen in the next activity.
Ramp Stimulus
One commonly used protocol to investigate the strength-duration relationship of a spiking neurone is to use a slowly-rising ramp stimulus, rather than a square-pulse stimulus.
There is a 20 ms, 1 nA stimulus applied, but instead of switching on instantly, the stimulus rises gradually to its final value over the Pulse 1Ramp duration. This fails to elicit an action potential.
Repeatedly click the up-arrow of the Pulse 1 stimulus Amplitude, until a spike occurs.
With each click, the stimulus strength increases by 1 nA. As before, the run-on-change and auto-clear options have been selected, so a separate experiment runs with each click. A spike should occur when the strength reaches 6 nA.
Refine the threshold amplitude value by reducing it in steps of 0.1 nA until the spike fails. Then increase by 0.1 again, to get a threshold value which can be compared to that of a square-wave 20 ms pulse in the original experiment.
Question: Why is the spike threshold higher when a ramp stimulus is applied compared to a square-wave pulse?
Refractory Period
The HH model provides a plausible mechanism for several well known properties of neurons, including their refractory period.
Note that two identical stimulus pulses are being delivered to the neuron, but that only the first elicits a spike. This is because the second stimulus occurs within the refactory period of the first spike.
Just to be sure that there is not something weird about the second stimulus, set the amplitude of the first stimulus (Pulse 1) to 0 and click Start.
Yes, the second stimulus is fine (I hope) and elicits a spike perfectly well. So it is definitely the presence of the first spike that prevents the occurrence of the second.
Return Pulse 1 amplitude to 50 (or reload the file).
Can we recover a second spike with a bigger stimulus?
Check the Run on change box in the Results view.
Click the up spin button of the Pulse 2 Amp to increase its value to 70.
The spin button delta was pre-set to 20 in the parameter file. You can check or alter this by right-clicking the button.
The second stimulus now elicits a spike! Clearly, the second stimulus is within the relative, not absolute, refractory period.
Next reduce the delay to the Pulse 2 by 2 ms (i.e. from 12 ms to 10 ms) by clicking its down spin button.
You are now back in the refractory period!
Increase the Pulse 2 amplitude in steps of 20 by clicking the up spin button until you get a spike, and then reduce the delay again by a step of 2 to 8 ms.
Repeat this for one more time, to find a spike at 6 ms delay.
At this point the spike you elicit is quite small (smaller than the voltage change produced directly by the stimulus itself) and the stimulus is so large that it would probably cook a real neuron in a real experiment. So in terms of real physiology, we have probably arrived in the absolute refractory period.
Question: Can you explain the refractory period in terms of the HH mechanism? To help with this load and Start the parameter file Refractory Details, and examine the relationship between the conductance traces (third axis) and gate probability traces (bottom axis).
Bonus Question: Spikes that occur in the relative refractory period get progressively smaller the closer they get to the first spike. Why? We will see an interesting consequence of this phenomenon when we look at the propagation of two spikes later in the tutorial.
Threshold Accommodation
It doesn’t actually take a spike to make a neuron refractory, although the name is usually only used for the spike-induced phenomenon.
A standard spike is initiated in response to the brief but quite large depolarizing Pulse 2 stimulus.
Set the amplitude of Pulse 1 to 2.
Click Start.
There is now a small but long duration depolarizing stimulus delivered before the brief pulse, and the latter now fails to initiate a spike.
Take-home message: Sub-threshold depolarization can have an inhibitory effect on subsequent stimuli that would otherwise have initiated a spike. This is called spike threshold accommodation, but its underlying mechanism is similar to that which causes the refractory period.
The stimulus current is now negative, and, perhaps surprisingly, this strong inhibitory stimulus produces a spike on “rebound”. Such rebound excitation is quite common in the nervous system, and frequently occurs after strong inhibitory synaptic input. What causes it?
In the resting axon, we tend to think of all the voltage-dependent channels as being shut. However, remember that while the opening and closing of channels is voltage-dependent, it is also probabilistic. This means that even at the resting potential, there will actually be a random number of potassium channels that are open (all their n-gates are open). Furthermore, for the same reason, there will be a number of sodium channels that are inactivated at rest (their h-gates are shut). This can be seen from the initial values of the activation variable probabilities in the lower trace. If we inhibit the neuron with strong negative current, then we will tend to close these open potassium channels (by reducing the value of n),
and remove the inactivation of sodium channels (by increasing the value of h).
The channel gates n and h have relatively long time constants at the resting potential,
and so these changes persist for a while after release from the hyperpolarizing stimulus.
This increases the excitability of the neuron, which can be enough to generate a rebound spikeThere may be other ion channels contributing to rebound excitation in other types of neurons. But the fact that it occurs in the HH model tells us that it is possible to get rebound with just the standard sodium and potassium channels..
Task: Use the simulation to obtain evidence for the mechanism proposed above.
Hint: Find the time at which the membrane potential reaches (and crosses) the resting potential after termination of the stimulus during the build-up to the
rebound spike, and look at the values of the underlying variables (m, h, n, the sodium and potassium conductances) at this time.
Compare these to their values before the stimulus, when the membrane potential has the same value (the resting potential),
but has not previously been subjected to a negative stimulus.
In one case (pre-stimulus) the membrane potential is stable, but in the other (during rebound) it is not.
The reason for the instability must lie in differences in the underlying variables.
Possible procedure:
Place a horizontal cursor at the resting potential, i.e. the membrane potential before the stimulus (-70 mV).
Zoom into the important part of the Results view by clicking the expand timebase toolbar button , and setting the upper scale of the conductance axis to 1.
Check the Measure box in the Results view.
Check the 2 cursors box in the Measure dialog, then check the Only difference box (which is now enabled).
Drag the blue cursor to just before the start of the stimulus (about 0.2 ms), where the membrane potential is at its resting level.
Drag the red cursor to the time where the horizontal cursor placed earlier crosses the membrane potential as it rises towards the spike (about 5.1 ms).
Click the Measure button in the Measure dialog.
The numbers in the Measure box show the difference between the before-stimulus values of various parameters,
and the values of those parameters at the same membrane potential (or almost the same) during the rebound.
To visualize these numbers place additional horizontal cursors in the Results view conductance and activation axes and
line them up with the pre-stimulus trace values. Note how much (if at all) the traces deviate from the horizontal cursors at the time of the red cursor.
Question: Which of the two effects mentioned above (potassium channel deactivation and removal of sodium channel inactivation) is mainly responsible for driving depolarization above the resting potential during rebound? Does the other effect play any role in the rebound mechanism?
Post-Inhibitory Facilitation
To make this a little more realistic, we can see rebound resulting from an inhibitory synapse.
This uses the Network model to simulate synaptic interactions between three different neurons, visible as yellow blobs in the main Setup view. The three neurons implement full Hodgkin-Huxley-type spikes. N1 makes an inhibitory synaptic input to N3, represented by the light-blue diamond labelled b, while N2 makes excitatory input to N3, indicated by the diamond labelled a. N1 and N2 both receive experimental stimuli, indicated by the square boxes above each neuron. The parameters of these stimuli are controlled in the Experimental control panel on the left of the main view. Both stimuli inject positive current and make the receiving neuron spike, but their delays are different.
The stimulus to N1 occurs quite early, and the N1 spike generates a hyperpolarizing IPSP early in the N3 trace (lower trace, blue). The stimulus to N2 occurs later, and its spike generates an excitatory input to N3, visible as an EPSP occurring after the IPSP. The EPSP is close to threshold, but does not quite make N3 spike.
In the Experimental control panel, note that stimulus 1 is selected in the stimulus list (if it is not, click it to select it). Now click the Delay up spin arrow to increase the Delay of Stimulus 1.
This brings the IPSP a bit closer to the EPSP, and the EPSP increases in size!
Click the up spin arrow several more times, observing the changes in the EPSP.
When the delay reaches 6 ms, the EPSP triggers a spike in N3.
To check that it really is the IPSP that is doing the job, Clear the screen and then check the strychnine box in the Setup view Drugs frame.
Strychnine blocks glycinergic inhibition (which is the transmitter mediating the IPSP). In the absence of the IPSP, the EPSP no longer generates a spike.
Uncheck the strychnine box, and increase the Stimulus 1 Delay by clicking the up spin arrow a few more times.
Note that as the IPSP gets closer to the EPSP, the facilitatory effect goes away, and the EPSP no longer initiates a spike. Eventually, the EPSP itself is diminished in amplitude as the IPSP negatively summates with the EPSP.
Take-home message: A correctly-timed IPSP preceding an EPSP can enhance the effectiveness of the EPSP.
Of course, it is important to remember that it is not the EPSP itself which is being affected here, it is the voltage-dependent channels that generate the spike which are affected by the preceding hyperpolarization. So it is the spike threshold of the post-synaptic neuron which is changing.
Exactly this effect has been shown to occur in auditory processing in the mammalian lateral superior olive (Beiderbeck et al., 2018). The inhibition in the circuit comes from the neurons activated via the Calyx of Held synapse, which has the distinction of being the largest known synapse in the mammalian brain – which presumably implies that it is important.
Pacemaker (continuous regular spiking)
The standard Hodgkin-Huxley model generates a neuron that only spikes if it is stimulated, either directly by a depolarizing stimulus, or indirectly on rebound from a hyperpolarizing stimulus. Indeed, since the squid giant axon on which the model is based mediates a sudden escape response, it would be highly non-adaptive if it spiked spontaneously. However, a small change in properties can result in a neuron that spikes spontaneously and repetitively, a type of activity known as pacemaker activity.
This is a standard HH model. The upper axis shows the membrane potential, the lower axis shows the conductance of the sodium and potassium channels. However, there is no stimulus, and absolutely nothing happens during the simulation run.
Reduce the maximum K conductance in the Membrane Properties frame of the Setup view by clicking the down arrow of the adjacent spin button.
This reduces the maximum possible potassium conductance (the conductance when all potassium channels are open) by 2.5 units. Effectively, it is as if the number of voltage-dependent potassium channels in the membrane had been reduced.
A new simulation runs automatically when you change the conductance (the Options: Run on change menu command has been pre-selected), but there is no obvious change in the output.
Reduce the maximum potassium conductance progressively by repeatedly clicking the down spin button.
When the conductance reaches 21 mS cm-2 the neuron starts to generate spontaneous repetitive spikes. The frequency of these spikes increases as you progressively reduce the maximum potassium conductance further. Note that the peak potassium conductance (the blue trace in the lower axis) becomes smaller with each reduction in the maximum. Eventually, the spikes become distorted in shape, and finally fail when the maximum potassium conductance reaches just 1.
Question: Why does reducing the number of voltage-dependent potassium channels lead to repetitive spontaneous spiking? Why do the spikes eventually fail when the potassium conductance is very low?
Spike Propagation (Conduction)
In the experiments so far, you have been using the standard HH model which simulates a space clamped axon. This means that the cell is isopotential (same voltage throughout) and the spike occurs simultaneously at all points across its surface – it does not propagate along its length. This is exactly the situation that Hodgkin and Huxley set up in their original experiments.
You are now going to look at the conduction of the action potential as opposed to its generation.
The Setup view shows a snaky string of blue blobs. This represents a non-myelinated axon, with each blob being a separate short sequential section of the axon (this is in fact a compartment model of the axon, and there are 50 compartments (blobs) in total). The blobs are colour-coded for membrane potential, where darker colours represent hyperpolarized potentials. The axon is initially at rest, hence the dark blue colour. The square 1 attached to the top-left blob (A1) represents a stimulus that will be applied to that end of the axon.
Click Start, and just watch the Setup view to begin with.
Re-run the experiment, but this time look at the Results view.
There are 3 recording traces, from electrodes placed at the start, middle and end of the axon (A1, A26 and A50 respectively). You see a spike in each trace and the spikes are virtually identical in height, but there is a time delay. This reflects both the all-or-none nature of the spike, and the fact that spikes have a finite conduction velocity.
Take-home message: Spikes propagate along an axon with delay but without attenuation (in contrast to passive propagation). This is why spikes are essential for long distance communication, since with purely passive propagation the signal would attenuate to zero within a few millimetres in most axons.
Click Clear.
Either
Drag the square stimulus 1 box onto the bottom blob. This will move the stimulus to the other end of the axon.
Or
Click the square stimulus box on the top blob to select it.
In the Experimental Control panel, change the Target neuron from 1 to 50.
Note that the stimulus box re-locates to the bottom blob.
Also note that although it’s called the target “neuron”, in this situation it is actually the target compartment within the neuron
Click Start, and note that the spike propagates perfectly happily in the other direction.
Take-home message: Axons are not polarized - they can conduct in either direction. The normal direction of propagation depends entirely on where the input stimulus occurs.
Now drag the stimulus to the middle of the axon (A26) and click Start.
You can even stimulate an axon in the middle, and spikes spread out in either direction. This is unusual, but not unknown, in real neural circuits.
Spike Collision
Reload the file Axon to get back to the starting conditions.
Either
Hold down the control key and drag the stimulus 1 box onto the bottom blob. This will duplicate the stimulus.
Or
Add 50 to the Target neuron(s) list in the Experimental Control panel (so that it reads 1 50).
(You will have to drag the new stimulus box to one side in order to see it clearly, because it will initially be superimposed on the A50 label.)
Click Start.
You are now applying a stimulus at both ends of the axon simultaneously. Spikes start to propagate towards the centre, and then they stop.
Question: What happens and why?
Spike Spatial Distribution
Reload the file Axon to get back to the starting condition
In the Results view, select Voltage vs Neuron from the Display mode options.
The Results view changes. It now shows a graph of membrane potential (Y axis) plotted against spatial location along the axon (X axis). Each of the 50 blobs shows its own membrane potential, at its own relative position along the axon.
Click Start, and watch the Results view.
Clear the screen and click Start again, but this time click Pause (the same button as Start) when the spike has reached about half way across the screen.
You can now see the voltage profile along the whole axon, at a time when the peak of the spike is located at about the mid point. In the Setup view you see the colours reflecting the membrane potential at each point. In the Results view you see a more detailed graphical display. There is a small pre-potential just in advance of the spike (to the right of the peak) which reflects the local circuit propagating in front of the spike wavefront. On the left of the view, you see that a substantial portion of the axon through which the spike has already passed (i.e. to the left of the peak) is hyperpolarized. This part of the axon is refractory.
Click Continue (the same button as Start and Pause) to allow the spike to continue to the end of the axon.
Passive vs Active Conduction: a qualitative comparison
At this point a quick review of the key differences between passive and active conduction might be useful.
A low amplitude but long duration pulse of negative current (-2.5 nA) is injected into the end of the axon. This spreads into the nearby adjacent sections of the axon, but gets weaker as it spreads. It is undetectable at section A20. This is visible both in the voltage vs time recordings in the Results view, and in the colour coding of the axon segments in the Setup view.
Click the down spin button arrow for the stimulus amplitude in the Experimental Control panel, to change the stimulus to -5 nA.
A new sweep starts immediately because Run on change is pre-selected.
Wait until the sweep finishes, and the click the down button again.
Repeat this one more time so that the amplitude becomes -10 nA.
Click the up spin button arrow for the stimulus amplitude, to change the stimulus to +5 nA.
So far, the signal (i.e. the current stimulus) is spreading within the axon through passive conduction. We have already looked at this in detail in previous tutorials, but here is a quick summary of some important features:
The signal attenuates (gets weaker) as it spreads - it does not get more than about a quarter of the way along this axon (and this is very short axon in terms of real biology).
The response is sign neutral - both positive and negative signals propagate.
The response to the signal is amplitude graded - the bigger the signal, the bigger the response.
A corollary of this is that, apart from the short delay as the response builds up, the strength of the
signal is directly coded in the strength of the response. It can be "read" from the immediate value of the membrane potential.
The response integrates (sums) the input signal - e.g. a brief negative stimulus imposed in the middle of the positive stimulus would produce a downward blip in the response. This is not shown in this simulation, but is included in this list for completeness sake.
What happens if we make the signal even more positive?
Click the up spin button arrow for the stimulus amplitude, to change the stimulus to +7.5 nA.
Now something radically different happens. The signal is still a long duration positive current, but the response includes twospikes, which are superimposed on the passive response at A1. And crucially, these spikes spread along the axon without losing amplitude. Both are visible at A20, at which point the passive signal has virtually disappeared, and one of them is visible at A48, although the other does not reach that point in the axon within the duration of the simulation (the colour coding in the Setup view shows that the spike is still located at about A35 when the simulation terminates).
Click the up spin button arrow again to change the stimulus to +10 nA.
Now the stimulus generates three spikes at A1.
To make sure that you are clear about the sequence, in the Results view set the Highlight sweep value to 8, to emphasise the last sweep in the series.
The spikes are propagating through active conduction. Here is a quick summary of some important features:
Spikes propagate with little or no change in amplitude, no matter how long the axon - i.e. they do not attenuate as they spread. This is the famous "all-or-none" rule in action.
Spikes have a finite conduction velocity - it takes time for a spike to propagate from one end of the axon to the other.
Spikes are only generated in response to positive signals that are above a threshold value.
The strength of the signal is coded in the frequency of the spike response - i.e. the number of spikes generated within a set time period.
A corollary of this is that decoding the strength of
the signal requires integrating (essentially counting) the number of spikes occurring over time. A single individual spike gives no indication about the stength of the signal that caused it, other than that it was above threshold.
Refractory Conduction
We saw earlier that in a space-clamped neuron, a spike that is initiated within the relative refractory period of a preceding spike, is reduced in amplitude. What happens if we look at the same thing but allow spike propagation?
This is a similar compartmental model to the Axon model we have just seen, but with a longer axon.
Note that there are 2 stimuli boxes attached to the top end of the axon.
Click Start.
There are two stimuli in rapid succession and they each generate a spike. You can watch the Setup view to see the progress of two spikes along the axon.
Once the simulation has completed, look at the top trace in the Results view. This is recorded from the start of the axon, at the site where stimulation occurs. Note that the second spike is smaller than the first because it is occurring in the relative refractory period of the first, just as we saw previously.
Now look at the bottom trace, recorded from the other end of the axon. The two spikes are now virtually identical in height, and there is a noticeably larger gap between them. To convince yourself of this (if you wish):
Insert two vertical cursors and position them over the spike peaks in the top trace.
Couple the cursors by selecting the View: Cursors: Vertical: Couple menu option.
Drag the left cursor so that it is over the first peak in the bottom trace.
Note that the right cursor moves with it, and illustrates the increased inter-spike interval.
Remove the cursors by right-clicking the Results view and selecting Del all vert cursors from the context menu.
You can also see this the effect by viewing the voltage profile along the whole axon.
Clear the screen, and switch the Results Display mode to Voltage vs Neuron as you did before.
Click Start, but as soon as the second spike appears, click Pause.
Click Continue, and allow the simulation to run to completion while watching the Results screen.
As the simulation progresses, the second spike gets progressively larger, but it also lags progressively further behind the first.
So what is going on?
At the start the second spike has to propagate by spreading a local circuit into a region of axon that is already partially refractory from the preceding first spike. This local circuit is thus acting on an “inhibited” region of axon, and it takes longer to drive the membrane above threshold. Thus the conduction velocity of the second spike is reduced. This means that it drops a bit further behind the first spike, and it is now in a less refractory region of axon. So the inhibitory effect is reduced, and the second spike gets larger, and starts to propagate a bit faster, but still slower than the first. This continues until the second spike has completely escaped from the refractory period of the first, at which point it is equal in amplitude, and propagates at the same velocity.
This is a rather advanced concept, and will not be found in many textbooks, but is clearly apparent in the model, and can be easily demonstrated in a real axon. It does not mean that refractoriness is irrelevant to circuit function – it still takes a bigger stimulus to initiate the spike in the refractory period (see the stimulus amplitudes), and that could be significant in a real circuit. However, it does mean that two spikes that start off close together with the second one in the refractory period of the first, will have separated out somewhat by the time they reach the other end of their axon, and the second spike will have grown in amplitude. This may have important consequences on the output of the neuron for phenomena such as synaptic summation and facilitation (although so far as I know this has not been investigated).
Myelination and Saltatory Conduction
Myelination is a well-known specialization that vertebrates (and some invertebrates) have evolvedThere is an interesting story that links myelination to the evolution of hinged jaws in the armour-plated placoderm fish early in the Silurian period (Zalc et al., 2008). to speed up axonal conduction. A myelinated axon is wrapped by multiple layers of glia cell membrane, which both reduces the effective membrane capacitance and increases the membrane resistance, thus increasing the length constant of the axon compared to what it would be without the wrapping, and possibly reducing the time constant as well (depending on the RC product). However, the wrapping is not continuous, but is interrupted at regular intervals spaced a few millimeters apart – the Nodes of Ranvier.
Voltage-dependent channels only occur at the nodes - the internodal membrane under the myelin sheath is purely passive. Thus action potentials are generated at the nodes, but then spread through passive conduction along the internode to the next node. During this spread the signal attenuates as normal for passive conduction, but due to the myelin the attenuation is less than it would be in an unmyelinated axon, and the potential is still above threshold when it reaches the next node. At this node a new spike is initiated, and the signal strength is restored. The action potential thus jumps from node to node (hence saltatory conduction from the Latin saltare, to jump), and the resulting conduction velocity is about 10 times faster than it would be in an unmyelinated axon of equivalent diameter.
As before, the Setup view shows a snaky string of blue blobs (compartments) representing the axon. However, only the compartments at the ends of the rows contain voltage-dependent (HH-type) channels and only these can generate spikes. These are the nodal compartments. The internode compartments under the the myelin sheath (represented by the pale purple rounded rectangles) are non-spiking. The internode compartments have both their membrane capacitance and leakage conductance reduced by a factor of 10 compared to the nodes, to take account of the insulating properties of myelin and its thickness. Note that this is not to scale – in a real myelinated axon the nodes are only 1 - 2 \(\mu\)m long, and the internodes may be several mm.
Click Start (you may need to set a slow-down factor to observe the effects more easily).
The Results view shows the membrane potential along the whole length of the axon simultaneously.
A depolarizing stimulus initiates a spike starting at the top-left of the axon. This spreads through passive conduction, and hence with attenuation, to the node at the right-hand end of the top row, where it again generates an action potential. This spreads passively to the left-hand end of the second row, and so on. Note that the spike in the top-right node also spreads back to the original top-left node, but this is refractory and does not spike. Hence the propagation is uni-directional once initiated.
Clear the Results view.
Set the Max time in the Trigger mode group in the Results view to 7 ms.
Click Start.
The simulation now stops after 7 ms, and which point the spike is centered at N21, which is a nodal compartment. (But note that membrane potential values are only calculated in compartment-wide steps, so the actual spatial distribution of the spike shape is distorted to give an apparent sharp rather than rounded peak to the spike.)
The colour map shows passive propagation of the depolarization both forwards along the axon (towards N26) and backwards (towards N16). This is confimed in the voltage vs distance display in the Results view (hover the mouse and look in the status bar to see where N26 and N16 occur in the display on either side of the voltage peak at N21). The voltage profile in advance of the spike (to the right of the voltage peak) is simple, because this is a region of unstimulated axon. However, the voltage profile further back in the axon (towards the left in the Results display) is complex. This is because it contains both spiking nodes which are currently in the afterhyperpolarization phase, and non-spiking nodes which are relaxing back towards their resting potential of -70 mV.
Clear the Results view.
Select Neuron vs Time as the Display mode in the Results view.
Click Start.
The Results view now shows a normal voltage vs time recording from 4 compartments along the axon – N1, N21, N26 and N31 (plus the N1 stimulus in the bottom trace). These compartments are each marked with a purple asterix in the Setup view for clarity.
N21 and N31 are nodal compartments (as is N1), and these generate normal spikes. However, N26 is in the internodal region, and its membrane is non-spiking. The signal in N26 is a result of passive propagation from the adjacent spiking nodes N21 and N31.
Question: Why are there 2 peaks in the signal in N26 (located in the middle of the internode in the middle row)? Hint: you may want to use a vertical cursor to examine the timing of these peaks relative to the spikes in the adjacent nodes.
Finally, note that this is a concept simulation that illustrates the mechanism of saltatory conduction, but it cannot be used to quantitatively compare conduction velocity in myelinated and non-myelinated axons. As stated above, the scaling is not accurate, and the values assigned to internodal membrane capacitance and leakage conductance are not based on quantitative evidence. Furthermore, the action potential kinetics in the nodal compartments are those of the HH model of the squid giant axon, which is not myelinated!
Voltage Clamp
Theory of the Voltage Clamp
A major experimental challenge in investigating spike mechanisms is to analyse the properties of the conductances that are responsible for spike production. The problem is that the conductances are voltage-dependent, i.e. they change as the voltage changes. This produces a circular chain of cause-and-effect: the conductance affects the current, which in turn affects the voltage, which then affects the conductance, and so on.
The voltage clamp is a piece of electronic apparatus that breaks this loop by fixing the voltage at a constant value set by the experimenter. Thus the user sets a desired membrane potential. The voltage clamp detects the actual membrane potential, and then very rapidly injects current into the neuron to correct any deviation of the actual potential from the desired potenials. The end result is that the voltage clamp injects enough current into the neuron to ensure that the actual membrane potential is always equal to the desired membrane potential set by the user. This happens very quickly (on a sub-millisecond timescale) if the voltage clamp is operating properly.
What is the point of this? When the voltage clamp shifts the membrane potential to the level set by the user, the voltage-dependent ion channels of the membrane adjust themselves to conductance values appropriate to that potential. Thus a depolarization will open sodium channels etc. Ionic currents flow through the conductances as usual and tend to perturb the membrane potential. However, the voltage clamp detects this incipient deviation from the user-set value, and immediately injects current across the membrane to counteract it. The amount of current injected is an exact mirror image of the current flowing through the ion channels that was trying to cause the change, and so by monitoring this current, the experimenter knows how much current is flowing through the ion channels.
Key points:
Any deviation of the actual membrane potential from the user-set desired value is caused by the flow of current through the ion channels, and so the current that has to be injected to stop the voltage changing is exactly equal and opposite to the ionic current.
Since the voltage is not changing (after the initial almost instant switch to the new potential), there is no current flowing into the membrane capacitance (see the capacitance equation in Basic RC Properties), and the experimenter can be confident that all the current is flowing through the ion channel conductors.
Simplified voltage clamp.
The diagram shows separate electrodes for measuring voltage and injecting current, which historically is the way the clamp was first implemented. However, most laboratories now use a single electrode voltage clamp, either continuous (cSEVC) or discontinuous (switched: dSEVC).
Thus in a voltage clamp experiment the important measurement is not the membrane potential, because this is set by the user, but rather it is the current that the voltage clamp produces in order to keep the membrane potential at this user-set level. The current generated by the voltage clamp is a mirror image of the current caused by the ions flowing through channels across the membrane.
There is usually a fairly simple relationship between ionic current flowing through a channel (I), the channel conductance (g), the membrane potential (Vm) and the reversal (equilibrium) potential (Veq) of the ion in question:
(We have already seen this equation in relation to the driving force, but it is important so it is repeated here for clarity.)
Thus if we know the membrane potential (set by the voltage clamp), the ionic current (measured by the voltage clamp) and the equilibrium potential of the ion in question (and that can usually be measured in the voltage clamp experiment itself, as we will see later), we can use this equation to calculate the ionic conductance at the clamp voltage.
The voltage clamp can also be used to investigate synaptic mechanisms. In this case, the change in conductance is not (usually) caused by a change in membrane potential, but rather by the opening of channels due to transmitter action. This is explored in a later activity.
There are actually 3 different types of currents that can be detected in a real voltage clamp experiment:
Ionic. The ionic current is as described above. It is what most experimental neuroscientists are interested in, and it is the only current that is actually simulated in the voltage clamp mode of Neurosim.
Leakage. This is just a subset of the ionic current, but it is the specific name given to current flowing through the leakage channels, as opposed to that flowing through voltage-dependent or ligand-gated channels.
Capacitive. This is the current that is applied by the apparatus whenever the voltage clamp needs to induce a step change in membrane potential. It is a large but very brief pulse of current that charges the membrane capacitance, and a key aim of the technique is usually to keep this current separate from any subsequent ionic currents. It is ignored in Neurosim (beyond a simple demonstration shown later).
Gating. Gating currents are tiny currents caused by the movement of charged amino acids within the channel proteins themselves as they undergo the conformational changes associated with opening and closing. The discovery of these currents provided good support for the fundamental HH idea of there being gates within the channels that move in a voltage-dependent manner, although the detailed characteristics of the current show that the reality is more complex than the simple HH model. Gating currents are not included in any simulation in Neurosim.
This shows a classic voltage clamp record of the type carried out by Hodgkin and Huxley. The upper trace shows the user-determined membrane potential. Initially the voltage clamp keeps this at the holding potential, which in this case is set at the resting potential (-70 mV). The voltage clamp then suddenly shifts the membrane potential to a new value, the clamp potential, which in this case is 0 mV. The membrane is held at the clamp potential for the period of the clamp duration, and then returned to the holding potential. Remember, all this is specified by the scientist as the desired experimental protocol.
The current that the voltage clamp has to inject to keep the membrane at these various potentials is measured, and displayed on the lower trace. Thus at the holding potential, the clamp current is zero, because the membrane is at the potential that it “wants” to be at anyway, i.e. the resting potential. When the membrane potential is switched to 0 mV, there is an initial negative, downward deflection of the current trace, meaning that the voltage clamp was injecting negative current. This means that there must have been an inward flow of positive ions across the membrane (or possibly an outward flow of negative ions), since the clamp had to inject negative current in order to counteract the incipient depolarising voltage shift that this flow would have produced. This inward flow is followed by an outward flow of ions across the membrane, signalled by the positive, upward deflection of the clamp current trace needed to counteract the incipient hyperpolarising voltage shift. It is these changes in the voltage clamp current that the scientist would take as the results of the experiment.
[Important note: in this simulation the current trace only shows the ionic current, the capacitive current needed to actually bring the voltage to its desired level is not shown. We will come back to this later, so don’t worry if you don’t quite understand it yet.]
At the moment, the current trace is hard to interpret because the current is carried by a mixture of ions. However, there are potent drugs that can selectively block specific ion channels.
In the Setup view check the TTX box (or click the test tube icon).
Click Start.
The inward (downward) current has disappeared because TTX (tetrodotoxin) selectively blocks voltage-dependent sodium channels, so what is left is the outward potassium current.
Uncheck TTX and check TEA, then click Start.
TEA (tetraethyl ammonium ion) blocks voltage-dependent potassium channels, so now the outward current is gone, leaving just the inward sodium current.
Click Clear.
Check both the TTX and TEA boxes, then click Start.
There is still a very small step change in the current trace.
You can zoom in on this by dragging over it with the vertical magnifier ( ) from the Results toolbar, then return to the default scale with revert ().
Question: What causes this small current step? Hint: what conductance is left when both voltage-dependent channels are blocked?
Visualize currents independently
Clear the screen and uncheck any applied drug.
In the Results view, click the Traces button to open the Trace and Axis Setup dialog.
In the Current section, check the Na, K and Leak boxes and uncheck the Total box.
Click OK.
Click Start.
In the simulation we can colour code the various currents separately, which makes life much easier than in a real experiment.
Note that we are now only observing the sodium current: the potassium channels have been blocked with TEA.
[Actually, the Trace setup has been configured to only show the sodium current, so we are not seeing the leakage current either.]
There is a small depolarising clamp pulse (from a holding potential of -70 mV to a clamp potential of -50 mV),
and a small transient inward (downward) sodium current shown in the middle trace.
For clarity, the 0-current level is marked by a dashed grey line.
The bottom trace shows a small transient increase in the sodium conductance.
This conductance CANNOT be directly measured in a real experiment.
In the simulation it has been calculated using the driving force equation ,
and it is included to make explanations simpler.
We now want to progressively increase the voltage clamp potential.
Click the up spin button on the clamp 1 potential to change the value to -30.
The delta value of the spin button was pre-set to 20 in the parameter file. You can check this by right-clicking the spin button.
Options: Run on change has already been set in the parameter file.
Repeat until you reach a clamp potential of +70 mV.
At this stage, the Results view will look at bit of a mess, with a lot of traces superimposed on each other. However, we can clarify things easily:
Click the up arrow on the Hilight sweep control in the Results View.
The sweep showing the first experiment you ran is now highlighted, while the others are greyed out.
Click the Hilight sweep up arrow repeatedly, to view each sweep in turn.
As you compare the peak response at each clamp potential, you should notice that for the initial increments in potential the sodium current peak increases downwards at each step, but then it starts to decrease again. Eventually the current trace deflects upwards. Note that zero current is in the middle of the current axis, so the change from a downwards to an upwards deflection reflects a change from inward to outward current: i.e. a reversal in the direction of current flow.
If you look at how the current changes with time at any given clamp potential (i.e. changes in the horizontal direction of a single trace), you see that the current increases to a peak or trough reflecting activation of the sodium channel, but then, even though the voltage is held steady, the current declines back towards the central 0 position, reflecting inactivation of the channel.
Task: Plot an "I/V"graph to show the relationship between the clamp potential (X axis) and the peak (positive or negative) sodium current at that potential.
Set the Hilight sweep to 1.
Check the Measure box in the Results view, to show the Measure dialog.
Drag the vertical red measure cursor to align with the peak (maximum excursion, in this case a trough) of the sodium current.
Click the Measurebutton in the dialog, and note that the values appear in the display.
Set the Hilight sweep successively from 2 to 7, and repeat the measurement for each sweep, repositioning the measurement cursor as appropriate.
Note that you can use the < and > buttons in the dialog or the keyboard arrow keys to fine-position the measurement cursor.
Also note that the peak occurs earlier in each sweep . The sodium channels respond more quickly when they are more depolarized.
At this stage you could click Copy in the measurement dialog, and paste the data into the graphing program of your choice. But we can also produce a simple plot in Neurosim itself:
Click the Plot button in the dialog.
Select Voltage from the drop-down list for the X axis.
Select I Na from the drop-down list for the Y axis.
You should end up with a graph looking rather like this:
We can use this graph to find the equilibrium potential of sodium, and hence to calculate the conductance throughout each clamp potential step.
Sodium Equilibrium Potential
Remember the driving force equation? Click the link to remind yourself if necessary.
There are two non-exclusive circumstances in which the sodium current (INa) can be zero:
Either the sodium conductance (gNa) is zero,
or the membrane potential (Vm) is equal to the sodium equilibrium potential (Veq), so that the driving force (Vm - Veq) is zero.
Task: Identify the two zero-current positions on the graph. (Hint: one is explicitly visible, but the other you have to predict by extending the line a bit.)
You can change the X axis scale in the plot by unchecking Autoscale X and editing the axis values.
You can get the value at any screen location by hovering the mouse at that location, and looking at the value in the X or Y read-out at the left of the dialog.
Question: What is the sodium equilibrium potential? Can you see why the equilibrium potential of an ion is sometimes called the reversal potential?
Close the Plot, Clear the screen and click the Run on change checkbox in the Results view to deselect it.
Enter your estimate of the equilibrium potential as the Clamp 1 potential and click Start.
If you have got it right, the current trace should be flat, even though the conductance trace shows a big increase. Make sure you understand how that can be.
Task: Use the driving force equation \eqref{eq:eqDrivingForce} to calculate the peak sodium conductance during the +70 mV clamp step. Does it match the value shown in the bottom (conductance) trace in Neurosim?
If you have closed the Measurement dialog, reset the Clamp 1 potential to +70 and click Start to generate an outward sodium current. Then measure the current. If you have not closed the dialog, you can read the values from row 7.
A note on units: In the HH module of Neurosim, current and conductance values are scaled to a 1 cm2 patch of membrane. This matches what Hodgkin and Huxley did in their original papers. However, most neurons have a surface area which is considerably less than 1 cm2, so the actual values will be proportionally lower. In the Advanced HH module which we will see later, the units reflect those in the actual neuron.
Sodium Inactivation
The characteristics of sodium inactivation which contribute to the refractory period can be nicely demonstrated using the voltage clamp technique.
Two identical clamp pulses are separated by a short interval. The second pulse produces substantially less conductance change and inwards current than the first. (Remember, the current is the only thing that can be directly measured in an experiment, the conductance has to be calculated).
In the bottom axis we see the activation variables. These can never be determined in a real experiment – they are simply part of the model. But examination of their traces shows why the model successfully simulates inactivation, and it is likely that something like this is happening at a molecular level in a real neuron.
The m gate variable (red trace) increases rapidly at the onset of the clamp pulses, and decreases equally rapidly at their offset. The profile is essentially identical for the two pulses, so inactivation is nothing to do with the m gates.
The h gate variable (green trace) decreases during the first pulse, and then starts to recover after the pulse terminates. However, the rate of change is quite slow, and recovery is by no means complete when the second pulse starts. So the second pulse is applied to sodium channels that are substantially still inactivated (h gates shut) as result of the first pulse. So they cannot re-open and the overall conductance is less. (Rember, the gate variables are probabilities, so if the value of h is 0.5 for instance, it means that half the total population of sodium channels are inactivated.)
Increase the clamp Interval to 8 ms, and click Start.
The current and conductance responses to the second clamp pulse are much bigger. This is because the h gate variable has had more time to recover, so fewer sodium channels are inactivated at the start of the second pulse.
Use the Results Hilight sweep facility if you are confused by the superimposed traces.
This is very similar to the Sodium IV file used earlier, but now we are looking at potassium currents and conductance (and note the changed potassium current scales).
Run a series of voltage clamp experiments up to a clamp potential of +70 mV, just as you did previously.
Note that you don't need to use Highlight sweep to measure the traces individually -
just place the red cursor towards the end of the voltage clamp pulse and click Measure once in the Measure dialog.
The value of all 7 sweeps will be measured simultaneously at that time.
Take-home message: There are two important points from this set of experiments. The first is that the potassium current increases continuously as you increase the depolarised level of the clamp potential; there is no sign of a reversal in the direction of current flow. The second is that the potassium conductance increase is maintained for as long as the depolarisation is maintained; the potassium conductance does not inactivate.
Tail Currents
There is no command potential in the potassium I/V graph at which the direction of the potassium current reverse. Thus we can't find an equilibrium potential using the same method that we used for the sodium current.
The problem is that the potassium equilibrium potential is quite negative, and is in the voltage range in which the potassium channels are shut anyway due to their voltage-dependency. So we don’t know whether zero current is due to zero driving force or zero conductance (check \eqref{eq:eqDrivingForce} if you need a reminder).
However, we can get round this by taking advantage of the fact that potassium channels respond quite slowly to changes in voltage.
The top trace shows an initial large depolarizing step to + 20 mV. This increases potassium current (middle trace) as a result of increasing potassium conductance (lower trace – again remember that this cannot be measured directly in a real experiment). Note that the rate of change is relatively slow, but that by the end of the 10 ms step the conductance and current have stabilized at their new values.
At the end of the first step, instead of returning all the way to the holding potential of -70 mV, there is a second step to a reduced depolarized level of -20 mV.
At the time of the step down to the new voltage, there is an immediate drop in current (the trace drops vertically), which is followed by a more gradual decline. The latter gradual decline is called the “tail current”.
What is going on?
As soon as the voltage drops from +20 to -20 mV, there is an immediate drop in driving force, since the membrane potential is now closer to the (unknown) equilibrium potential. This causes an immediate drop in current (check \eqref{eq:eqDrivingForce}). At this point there has been essentially no change in conductance – it is still at the same value as it was at the more depolarized voltage because of the slow kinetics of the potassium channels. Then, as time passes, the potassium channels “catch up” with the reduced depolarization and start to close. This causes the gradually declining current that constitutes the tail current.
Task: At this point we actually have enough information to estimate the potassium equilibrium potential. We can construct two driving force equations \eqref{eq:eqDrivingForce}, one for the top of the vertical current step, one for the bottom. We know the numerical values of the membrane (clamp) potential (Vm) for each step (from the Setup view, or by measuring it in the Results view) and can measure the current (I) at the top and bottom of the vertical drop. The conductance (g) and equilibrium potential (Veq) is the same in both equations (the conductance is the same because it has not had time to change, and the equilibrium potential is the same because it is fixed by the Nernst equation). So by making g the subject we can eliminate it, and solve the remaining equation for Veq.
However, we can also just do some more experiments to find the equilibrium potential.
Set the Clamp 2 Potential to -100 mV and click Start.
The tail current has now reversed. The gradual change in current following the step drop is now in an upwards direction. So we have gone beyond the equilibrium potential.
Task: Make successive changes to the Clamp 2 Potential to try to eliminate the tail current completely. I.e. find a clamp potential where the current trace is completely flat after the vertical drop.
This is the equilibrium potential, where there is zero current because there is zero driving force with a non-zero conductance.
Check the Run on change and Auto clear boxes in the Results view to speed things up.
Repeatedly click the up-arrow of the Clamp 2 Potential, and note the changes in the tail current.
Take-home message: Tail currents are quite hard to understand. However, the mental effort involved is a really good way of getting to grips with some important concepts underpinning cellular electrophysiology.
Capacitive Current (voltage clamp)
A key requirement of voltage clamp experiments is that the voltage should change extremely quickly when it steps to a new value. This is to ensure that there is a complete separation between capacitive and ionic currents – all the capacitive current flows during the voltage change, meaning that during the period of stable voltage, any changes in clamp current reflect changes in ion flow across the membrane.
How does the voltage clamp get the membrane potential to its desired level so rapidly?
The answer is that when a voltage step change is required, the clamp apparatus produces a very large but very short-lived current pulse to rapidly charge the membrane capacitance, and hence rapidly change the voltage. This is called the capacitive currentAlso the capacity or capacitative current..
We can visualize this using a current clamp protocol.
For clarity, this neuron has no voltage-dependent channels, it just has fixed-conductance leakage channels. A pulse of current is injected (stimulus box 1 attached to the neuron in the Setup view), and the membrane rises with an exponential curve towards its final voltage value, as expected for the standard RC passive properties. The resting potential is -60 mV, and the final voltage is about +4 mV.
A reminder: during the rising phase of the voltage, part of the stimulus current is going into the membrane capacitance, and part through the leakage channels. However, by the time the voltage stabilizes, all the current is going through the channels.
In the Experimental Control panel, click on Stimulus 2 in the stimulus list to select it (or click on stimulus box 2 in the Setup view)
Note that the initial amplitude of stimulus 2 is 0, i.e. the stimulus is silent.
Set the Amplitude of the Stimulus 2 to 4.4 (I found this value by trial-and-error).
Click Start.
The new sweep shows a large extra pulse of stimulus current at the start, and now the voltage rises almost instantaneously to its final value.
Click on Stimulus 3 to select it.
Set its Amplitude to -4.4 (symmetrically opposite to Stimulus 2).
Click Start.
We now have a symmetrically opposite large stimulus pulse at the end of the main stimulus, that brings the membrane rapidly back down to resting potential.
Use the Highlight sweep facility to observe each of the 3 conditions, leaving the third sweep highlighted at the end.
The important point illustrated in the third sweep is that during the main part of the stimulus, the voltage trace is flat (dV/dt = 0), which means that all the stimulus current is going through the ion channels, which in this case are just the leakage channels (see this equation). The current going into the membrane capacitance is limited to the large capacitive current at the start and end of the main pulse.
A voltage clamp basically does exactly this, but with a reversed control logic. The experimenter would set the desired clamp potential to 4 mV. The voltage clamp apparatus would detect that the membrane potential was actually at -60 mV, and would inject a large current pulse to rapidly shift it positive. It would start off extremely large (probably as big as the apparatus can manage), and then be reduced as the apparatus detected that the voltage was approaching the desired 4 mV level. Once the 4 mV level was reached, the apparatus would continue to inject just enough current to hold it there. If there are no voltage-dependent channels, then the amount of clamp current needed to hold the clamp potential steady would be a fixed constant value depending on the leakage driving force (as we see in this simulation). If voltage-dependent channels were present and started to open, the voltage clamp would detect the incipient deviation from 4 mV resulting from current flow through those channels, and would inject current to prevent the voltage from changing significantly.
Why a Constant Voltage Matters
While the voltage is changing, only a part of any clamp current is going through the ion channels, the other part is charging the capacitor. Ion channels can change their conductance, and hence the current flow through them, on a millisecond timescale. So if the clamp potential rises slowly, then during the changing voltage you cannot sort out what is capacitive current and what is ionic current – they are inextricably mixed together. By having a large capacitive current that is over almost instantly, we know that any current after the capacitive current, will be current needed to keep the voltage at its stable clamp potential, and this is current actually flowing through the ion channels.
If we know the moment-by-moment ionic current and we also know the equilibrium potential for the ion in question (which we can usually find with an appropriate voltage clamp experiment to find the I/V relationship), then we can use the driving force to calculate the moment-by-moment channel conductance. That is usually what we are interested in, since that knowledge may shed light on the mechanism underlying whatever cellular phenomenon we are investigating.
Space-Clamp Problems
One of the technical problems in real voltage clamp experiments is ensuring that the voltage is uniform throughout the whole cell, a condition known as a space clamp. If it is not uniform, then current can flow from the region of the cell where the voltage is different to the clamp potential into the region of the cell supposedly under voltage clamp, and contaminate the results. This is not a problem for spherical cells, which are naturally isopotentialIsopotential means that there is the same membrane potential at every point on the surface., but it is a problem for axons, or cells with an extensive dendritic field. Hodgkin and Huxley solved this by inserting a highly-conducting silver wire internally along the whole length of the axon they studied, which held the axon at a uniform potential, but this is rarely feasible. The best approach is to work with cells that are as small as possible so that axial flow is minimized. However, often there is no perfect solution and researchers just have to live with the contamination and try to argue that it is not sufficient to invalidate their conclusions.
Single Channel Patch Recording
Conductance
We have previously seen single channel patch recordings during normal, unclamped action potentials. We can also look at such recordings with a voltage clamp protocol, which is actually how single-channel recordings are normally made.
This shows a voltage clamp experiment with a long duration clamp step. There are traces showing the macro (whole cell) sodium and potassium conductance on the 3rd axis, and single channel conductances on the 4th and 5th axis. Note that the single channel conductance values are normalized - an open channel has a conductance of 1 (a closed channel has a conductance of 0, of course). Also, remember that you cannot directly measure conductance in an experiment, you can only measure current and then calculate conductance. But in a simulation we can get the computer to do the calculation for us ...
Click Start a few times to get a feeling for when the single channels open and close.
Then check the Gates cartoon box in the Result view.
The Membrane Channel Model window opens showing a visual representation of the HH model of sodium and potassium channels.
Move the window so it does not obscure the Results.
Select a Slow down factor of 8 from the drop-down list in the main toolbar. This will enable you to see the cartoon animation more easily. You may need to adjust this value depending on the speed of your computer.
Click Start, and observe how the individual gates in the cartoon open and shut.
When all the gates are open an arrow is drawn through the channel to show that it is now conducting. This will coincide with an open step in the patch traces in the Results.
Add the drugs TTX, TEA and Scorpion toxin in turn to the preparation, observing how the response changes with each drug.
Reload file VC Patch Conductance to get back to the starting conditions.
Micro and Macro Conductance
Increase the Patch channel Count to 50 and click Start.
Hold down the control key and click the autoscale button in the Results toolbar to rescale the axes.
Now the patch recordings should start to look like the overall conductance traces in terms of their shape.
Take-home message: The overall sodium and potassium conductance expressed by the whole neuron is made up of large number of fixed-size single channel micro-conductances, which open and close probabilistically. (You may have seen this message previously in the unclamped simulation...)
Current
We are now going to look at the current flowing through single channels. This is more realistic in the sense that you could actually make these measurements in a real experiment.
The traces, from top to bottom, show the voltage applied by the clamp (which is set by the experimenter), the sodium and potassium whole-cell currents, and the currents through single sodium and potassium channels.
Repeatedly click Start until you detect a current through the sodium channel in the patch recordings (ignore the potassium channel for now). This may take a few tries.
Measure the current through the sodium channel in its conducting state.
Change the voltage clamp successively to 0, +35 mV and +70 mV, repeating the measurement for the sodium channel in its conducting state at each voltage.
Plot the Na patch current (pA) (Y axis) against the clamp Voltage (mV) (X axis). (Make sure that you plot the patch current, not the whole membrane sodium current, I Na).
This is an I/V curve for sodium, but it looks very different to the J-shaped whole-cell curve you may have produced earlier.
Question: Why do the whole-cell and single-channel I/V curves for sodium have such different shapes?
Task: Determine the single-channel conductance of the sodium channel in the patch.
Hint: from Ohm's Law, conductance = current/voltage, so the slope of the graph (which is indeed current/voltage) gives the value of conductance;
in this case you do not have to worry about the equilibrium potential or driving force. Make sure you get the units right.
It is easy to get the slope of a line in a Neurosim plot: just check the Linear trendline box and read the slope value.
Take-home message: Since the single-channel I/V plot is a straight line, the individual open sodium channels have a single, fixed conductance across the whole voltage range.
The HH module explicitly implements the original Hodgkin-Huxley model that was developed using the squid giant axon as the experimental preparation. In this model the only voltage-dependent channels are the sodium and potassium channels. However, since publication of the original model, a very large literature has developed in which researchers extend the model to include other types of voltage-dependent channels. In Neurosim, this can be done in the Advanced HH module. Neurons developed in the module can also be incorporated into circuits using the Network module.
Endogenous Burster Neurons
So far we have been concerned only with neurons that are silent unless they get a stimulus. However, some neurons have spontaneous activity even when totally isolated from any synaptic input. One such class of neurons are called endogenous bursters. These, as the name suggests, produce rhythmic bursts of spikes.
The top trace shows the membrane potential, and the rhythmic bursts of spikes are very obvious.
The parameters for this model have been adapted from those of the mollusc neuron R15, which is a well-known endogenous burster, but similar endogenous bursters occur in other animals, including in the respiratory centre of mammals.
Look at the Channels frame in the Setup view. This lists the 4 types of voltage-dependent channel present in the neuron. The top two channels, Na(V) and K(V), are basically HH-type channels that generate spikes. It is the other two that give the neuron its bursting properties.
Click Clear.
Check the TTX and TEA boxes in the Drugs panel of the Setup view.
Click Start.
Clearly, the neuron doesn’t need spikes in order to produce a rhythm (although it would need spikes to propagate the signal to the end of its axon). The actual burster mechanism must lie in the remaining two channels.
The voltage-dependent calcium channel in this model is low threshold and non-inactivating, with rather slow kinetics. It has a non-zero conductance even at quite hyperpolarised membrane potentials, so there is always an inflow of calcium ions which will tend to depolarise the neuron. As the neuron depolarises, the calcium conductance increases because it is voltage-dependent, so that the calcium inflow also increases. This is regenerative just like the sodium channel of a normal spike, and it drives the membrane up to a depolarized level. This is accompanied by a substantial increase in intracellular calcium concentration (visible in the bottom trace).
The other channel is a calcium-dependent potassium channel. This is dependent on both voltage and intracellular calcium concentration, which means that in order for this channel to be open, the neuron has to be both depolarised, and have a relatively high concentration of free intracellular calcium.
This is exactly the situation caused by the calcium channels, so the potassium channels open and terminate the "spike" and repolarize the neuron.
Once the neuron is repolarized, the rate of calcium inflow reduces because the calcium conductance drops. Calcium homeostatic mechanisms now reduce the intracellular calcium concentration, which, in combination with the repolarization, shuts the potassium channels.
At this stage the neuron is back where it started, and the cycle repeats.
In the Drugs frame in the Setup view, check the box labelled apamin. Apamin is a drug derived from bee-sting venom (hence its name – the honey bee is Apis mellifera), and it is a specific blocker of calcium-dependent potassium channels.
Click Start to see what happens. Hopefully, it makes sense.
Phase Resetting
One test for the proposed mechanism is to experimentally perturb the timing of the rhythm.
Clear the screen and remove (uncheck) the TTX and TEA and apamin, then click Start.
We are back with the original rhythm.
Do not Clear the screen. Set the stimulus Pulse 1 amplitude to 1 (it was previously 0 and thus switched off), and click Start.
For the first half of the screen the bursts should exactly superimpose, but when the stimulus pulse arrives, it induces an extra burst. After this stimulus, the timing of subsequent bursts is reset. They occur at the same frequency as previously, but there has been a phase advance introduced by the extra burst.
With the traces superimposed like this it can be difficult to see what is happening, but the following might help:
In the Results view, set the Hilight sweep first to 1, and then to 2. Hopefully that makes things clearer.
This sort of phase resetting is a standard test to see whether a neuron participates in generating a rhythm, or merely follows a rhythm generated by some other part of the neural circuit. We will see it again when we deal with central pattern generates in simulations dealing with networks of neurons.
Inactivating Calcium Channels
Many examples of bursting activity that have been investigated results from oscillations in membrane calcium conductance. However, the details vary from animal to animal and cell to cell. In the previous example the calcium-activated potassium current was essential for burst termination, but another common mechanism is for the slow calcium current to self-terminate through inactivation.
This neuron again has the standard sodium and potassium channels, but the only other voltage-dependent channel is a calcium channel. As before, this has a low threshold, but unlike the previous calcium channel, this one has voltage-dependent inactivation as well as activation.
Check the TTX and TEA boxes in the Drugs panel of the Setup view.
There is no need to click Start or Clear between changes because Run on change and Auto clear have been pre-selected
As before, the sodium and potassium channels are only needed for generating spikes, the oscillation comes from the calcium channel. The calcium channel activates at rest potential and depolarizes the neuron. Since the channel is voltage-dependent the depolarization is regenerative and the membrane potential takes off in a spike-like manner. However, after a relatively long delay the channel inactivates, and the voltage drops again. The channel then de-inactivates, allowing the cycle to repeat.
Channel Kinetics
Double-click Ca (inactivating) in the channel list in the Setup view.
Or single click it to select it, then check the Properties box.
DON'T PANIC!
The Voltage-Dependent Channel Properties dialog opens. This has a lot of facilities, but we will ignore most of them. (Press F1 to view the on-line help if your curiosity gets the better of you.) The dialog is non-modal and can stay open while you do experiments.
First note that although the channel Description says that it is a calcium channel, the carrier ion is actually Unspecified. You only need to specify it as calcium if you want to model the intracellular calcium concentration, which in this case we do not (there are no calcium-dependent channels).
In the Gates listing, there are 2 gate types, activation and inactivation.
Click Activation in the gate listing to select it (it may already be selected).
Look at the kinetic listing, and note that Tau max is 40 ms. This essentially determines how rapidly the activation gate responds to changes in voltage.
Click Inactivation in the gate list to select it.
Note that Tau max is now 7500. It is this difference in kinetics which is one key to the oscillation.
Click Activation to re-select it.
Check the Show graph box in the Equations group about half way down the dialog on the right. The dialog expands to show a graph which by default shows the steady-state voltage-dependence of the activation and inactivation gates. Both show a sigmoid response curve, but with opposite slopes.
Locate the parameter V mid in the Gate kinetics frame, which should be visible near the bottom of the dialog box. It should read -30 at present. This determines the voltage at which the steady-state open probability is 50%.
Click the down spin button beside V midfive times in succession to reduce its value to -35 mV in steps of 1 mV.
Note that the activation curve (red line) in the graph shifts slightly to the left with each change, and that the oscillation frequency (visible in the Results) increases. The left shift in the graph means that the calcium channel activates at a slightly lower voltage, which means that the depolarizing drive is slightly stronger, hence the frequency shift.
Continue decreasing V mid until the oscillations stop, which should occur when it reaches a value of about -38 mV.
Now click the up spin button repeatedly. The oscillations return as the V mid value approaches its original value of -30 mV, but then fail again as V mid reaches about -22 mV.
The Channel Properties dialog allows the user to fine-tune kinetic parameters, and to explore the consequences of changing parameter values. It is extremely difficult (actually, in most cases completely impossible) to predict from first principles the consequences of changing particular parameter values. The only way to find out is to run the model and try things out.
Conditional Bursters
There is not always a binary divide in neuron types between bursters and non-bursters - some neurons can change their output mode depending on the overall excitation level. Such neurons have been found in the respiratory centre of the mammalian brain (Butera, et al., 1999).
Load the file Conditional burster. (This is a direct implementation of the published model, and is actually the same neuron that was built in the Build a neuron walk-through.)
The Setup view shows that the neuron has 3 voltage-dependent channel types. There are simplified versions of the standard HH potassium (1: K delayed rectifier) and sodium (2: Na fast) channels, plus a persistent sodium channel (3: Na persistent, commonly abbreviated as NaP). This latter differs from the standard sodium channel in 3 key respectsIf desired, these properties could be examined by checking the Properties box and comparing the two sodium channels, but this is not essential.: first it has a lower overall conductance, second it activates at a slightly less depolarized potential (i.e. it turns on more easily with a stimulus), and third, and most importantly, althought it does eventually inactivate, it inactivates much more slowlyThis is obviously why it is called the persistent sodium channel. than the normal HH sodium channel (the fast channel). It is the third channel that gives this neuron its interesting properties.
At zero and low stimulus levels the neuron is silent. As the stimulus increases, NaP starts to activate and the rate of depolarization slowly accelerates.
Eventually, the neuron depolarizes to the point that the fast sodium channels activate, and the neuron generates a series of spikes. However, NaP eventually inactivates and the burst terminates after a few seconds. The afterhyperpolarization of the final spike in the burst somewhat de-inactivates NaP, and then the depolarizing current simulus causes another burst. For a while the neuron is in regime where it acts as an endogenous burster.
Eventually the stimulus is strong enough to elicit spikes even when NaP is partially inactivated. At this point the neuron is in a regime where it exhibits tonic pacemaker spiking activity.
The important role of NaP can be demonstrated by blocking it.
In the absence of NaP the membrane potential depolarizes more linearly and at a lower level. This is because it does not depolarize enough to activate the fast Na channels, so the membrane is essentially passive in this voltage range.
Clear the Results.
Set the Amplitude of Pulse 1 to 0.08 nA.
Click Start.
With the higher stimulus, the neuron generates standard HH spikes.
Take-home message: The core mechanism generating endogenous bursting normally requires two factors. First, there must be a channel with a low activation threshold that mediates an inward current to drive the depolarizing phase. This could be either a sodium or calcium channel. Second there must be a slow-acting factor to terminate the depolarization with delay - after it has had time to initiate a burst of spikes. The factor could be simple slow inactivation of the depolarizing channel, or it could be activation of a potassium channel due to the accumulation of calcium entering during the depolarized phase.
Plateau Potentials
Some neurons show plateau properties. Such neurons are not endogenously active, so in the absence of a stimulus they are silent. However, a brief excitatory stimulus, such as a current pulse or EPSP, causes them to produce a long-duration multi-spike response that outlasts the stimulus. One neuron with such properties is in fact the respiratory neuron seen in the Conditional burster section above.
This neuron is exactly the same as that used in the previous section, but I will repeat its description here in case you have jumped straight into this section.
A very brief excitatory stimulus is applied, and this activates a HH spike in the normal way. However, it also activates the persistent sodium channel (NaP), and this remains activated, causing a sustained burst of spikes. This is the plateau potential.
Question: Why does the plateau potential finally terminate?
Click the down spin button of the Pulse 2 amplitude control in the Setup view to set a value of -0.2 nA.
Click Start.
The negative Pulse 2 stimulus now terminates the plateau potential before it would have naturally finished.
Note that in this model the timing of the inhibitory stimulus is quite important. If it occurs between spikes in the burst, as is the case here, then quite a small negative stimulus will terminate the burst, but if it occurs during a spike then it requires a much stronger negative stimulus to overcome the efffects of the inward currents driving the spike.
The effects of NaP can be seen more clearly by removing the standard HH currents.
Return Pulse 2 to 0.
Click Start to generate another full-length plateau potential.
Select 1: K delayed rectifier in the Setup view, and check the Block box.
Repeat for 2: Na fast.
Click Start.
You should now see a plateau potential without superimposed spikes. It differs from the normal plateau in two ways. First, it lasts longer - it is still activate at the end of the simulation run. Second, the plateau depolarization is much greater than with the HH channels active. These two differences indicate that activation of the voltage-dependent potassium channels normally restricts the overall depolarization and hence the level of activation of NaP. Without being "help back" by the potassium current, NaP achieves a higher level of activation, and takes longer to inactivate.
Clear the Results view.
Lengthen the simulation run-time by clicking the compress button in the Results toolbar ().
Alternatively, set the simulation end time to 2000 ms directly by editing the right-hand scale of the X axis.
Click Start.
The NaP-mediated plateau potential does terminate, it just takes longer than when HH spikes occur.
Uncheck the Auto clear box in the Results view.
Unblock the two HH channels, and run the simulation again to see the difference.
Note: In this particular neuron the plateau potential is generated by a persistent sodium current. However, very similar effects can occur in neurons containing slowly-inactivating (L-type) calcium channels.
Take-home message: Plateau potentials can be initiated by brief excitatory stimuli, and terminated by brief inhibitory stimuli. They are usually generated by slowly-inactivating sodium or calcium channels
Bi-stable neurons
Plateau potentials terminate because of eventual inactivation of their driving current, and so far as I am aware, in most real neurons plateau potentials do eventually terminate. However, in principle they could last indefinitely if the channel did not inactivate at all, thus producing a bi-stable neuron - one that was either inactive, or active, and that could be switched between states by brief stimulus currents or EPSPs and IPSPS. Such a neuron could form a single-cell flip-flop component in a neural circuit (you can see such a circuit in the Network section here).
Reload the parameter file Plateau to get to known starting conditions.
Click Start, and note the automatic termination of the plateau.
Double-click 3: Na persistent in the Channel list in the Setup view, to open the Voltage-dependent channel properties dialog.
Select h in the Gates list in the dialog.
Click the Del button to remove the inactivation gate from the channel.
Click OK to dismiss the dialog.
Click Start.
Now the spike burst persist for the full length of the simulation. It would, in fact, carry on indefinitely. Can such a burst be terminated?
Check the Run on change box in the Results view.
Click the down spin button of the Pulse 2 amplitude control in the Setup view to inject a pulse of -0.2 nA. This does not terminate the burst.
Click it again, to inject a pulse of -0.4 nA. Note that the burst now terminates.
It takes more negative current to terminate the burst in the bi-stable neuron than the plateau neuron because NaP no longer has any inactivation to aid the termination.
Take-home message: A bi-stable neuron can be switched between stable inactive (silent) and stable active (spiking) states by brief excitatory and inhibitory stimuli respectively.
'A' Current (Transient Potassium Channels)
Potassium channels in the standard HH model do not inactivate (because those in the squid giant axon that Hodgkin and Huxley studied do not). However, there are many different types of potassium channel, and some of them do inactivate. The most widespread of these belong to a family known as the A-type, or transient, or fast potassium channels. These channels are very common in neurons of the vertebrate nervous system, including the brain. However, the simulations in this part of the tutorial use the kinetic parameters published by Byrne (1980a, b) which come from a molluscan system, so the currents are all rather slow (sluggish?) compared to the vertebrate systems, but the general principles are the same.
Look at the Channels frame in the Setup view. It shows that the neuron contains three types of voltage-dependent channel: a standard sodium channel (Na (V)), a standard potassium channel (K (V)), and a fast potassium channel (K (A). The first two are essentially slow versions of the Hodgkin-Huxley channels (although the potassium channel is slightly different in that it does eventually inactivate with maintained depolarisation - but don’t worry about this for now). It is the third channel to which we will pay attention, the fast potassium channel K (A).
The Results view shows a spike generated in response to a depolarizing stimulus. What does inclusion of the A-type potassium channels do to the spike properties?
A drug called 4-aminopyridine (4-AP) specifically blocks the fast potassium current, so:
Check the 4-AP box in the Drugs panel of the Setup view
Click Start.
There are now two spikes, whereas before there was just one.
The effect of removing the A current with 4-AP has been to increase the rate at which spikes are produced in response to the stimulus.
When they are present, the A channels activate rapidly, and, being potassium channels, they oppose the depolarizing stimulus and slow down spike production.
However, the A channels inactivate, and after they inactivate, their negative influence is removed, and the neuron spikes as normal.
The dynamics of A-type channels can be appreciated more readily under voltage clamp.
Remove the 4-AP.
Click Clear.
Select the Voltage clamp mode.
Click Start.
The top axis shows a standard voltage-clamp protocol: hold at resting potential, step to 0 mV, then return to resting potential.
The middle axis shows the resulting currents. The total current, which is what would be recorded in a real experiment in the absence of drugs, is shown in the pale blue (cyan) trace. It looks very much like the classic HH voltage clamp response. However, judicious application of drugs could separate this out into 3 components, but to save you the bother this has been done programmatically (through the Traces button). The brown trace is the Na(V) current, and the green trace is the K(V) current. These are similar in form to the standard sodium and potassium currents of the HH model. The blue trace is the K(A) current, which is the one we are interested in.
The K(A) current activatesrapidly compared to K(V), which is why it is sometimes called the fast potassium current. Also note that it inactivates (the current declines even though the clamp depolarization is maintained), which is why it is also sometimes called the transient potassium current. (There is no obvious explanation as to why it is generally called the A current – my guess is that it was one of the first non-HH potassium currents discovered.)
Take-home message: The A-current delays the spiking of neurons in response to a stimulus because it is carried by potassium channels that activate early, but it does not prevent their spiking if the stimulus is maintained, because the channels eventually inactivate.
Inking in Aplysia
The following simulation is based on the work of Byrne elucidating the mechanisms underlying the inking behaviour of the marine mollusc Aplysia. This is a particularly satisfying story, because it is one of the few examples where a major aspect of the quantitative behaviour of a whole animal can be explained by the properties of a single ionic current (Byrne, 1979). The essential features of the story are as follows. When attacked by a predator, Aplysia may defend itself by releasing a "smokescreen" of cloudy ink, behind which it will attempt to escape. However, this is a rather expensive process for the animal (since it has to synthesise the ink), so it will only produce the behaviour in response to a very persistent stimulus. In the laboratory, inking can be elicited by directly stimulating sensory neurons, but again, the stimulus has to be maintained for 1 - 2 seconds. A brief stimulus fails to elicit inking.
The stimulus pulse is 200 ms in duration, and it fails to elicit spiking in the neuron. Note that the current and conductances traces only show the A-current response.
Increase the duration of stimulus pulse 1 to 800 ms.
Click Start without clearing the screen.
The neuron now spikes in the later part of the stimulus. The explanation for this "delayed reaction" is visible in the lower two traces. The A current activates as soon as an excitatory (depolarising) stimulus arrives. The A current then counteracts this stimulus, and prevents it from eliciting spikes. The A current eventually inactivates, and the long duration excitatory stimulus can now depolarise the neuron to threshold, and induce spiking and hence inking. In this way, the A current introduces a delay in the response at a monosynaptic excitatory synapse, and this delay is mirrored right through into the behaviour of the whole animal.
Apply the drug 4-AP.
In the absence of the A-current, the neuron now spikes immediately the stimulus is applied.


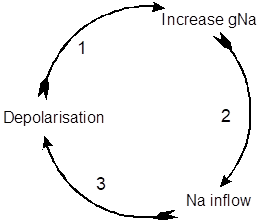
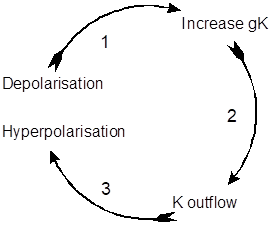
, vertical:
), or with options in the View: Cursors menu.
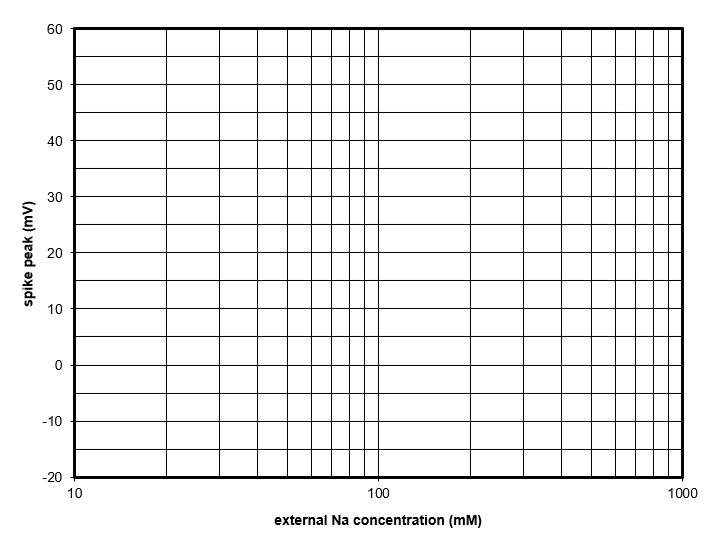

 in the Results toolbar to rescale the axes.
in the Results toolbar to rescale the axes.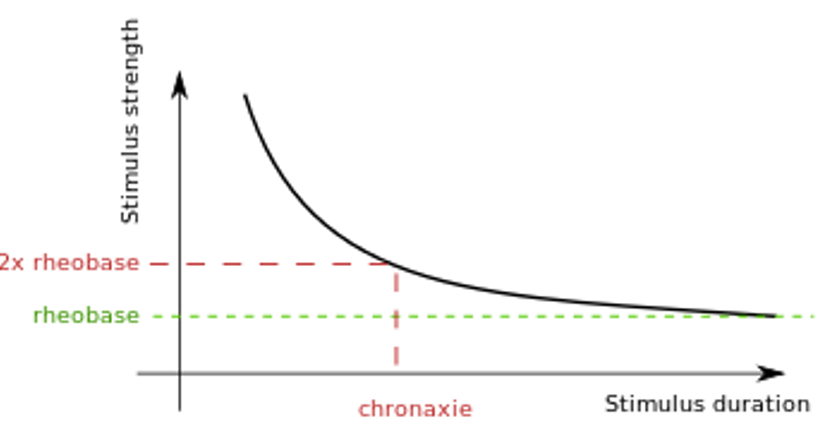
 ) in the Results toolbar.
) in the Results toolbar. , and setting the upper scale of the conductance axis to 1.
, and setting the upper scale of the conductance axis to 1. 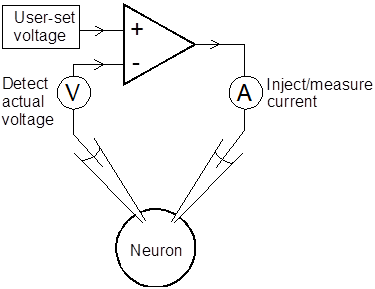
 ) from the Results toolbar, then return to the default scale with revert (
) from the Results toolbar, then return to the default scale with revert ( ).
).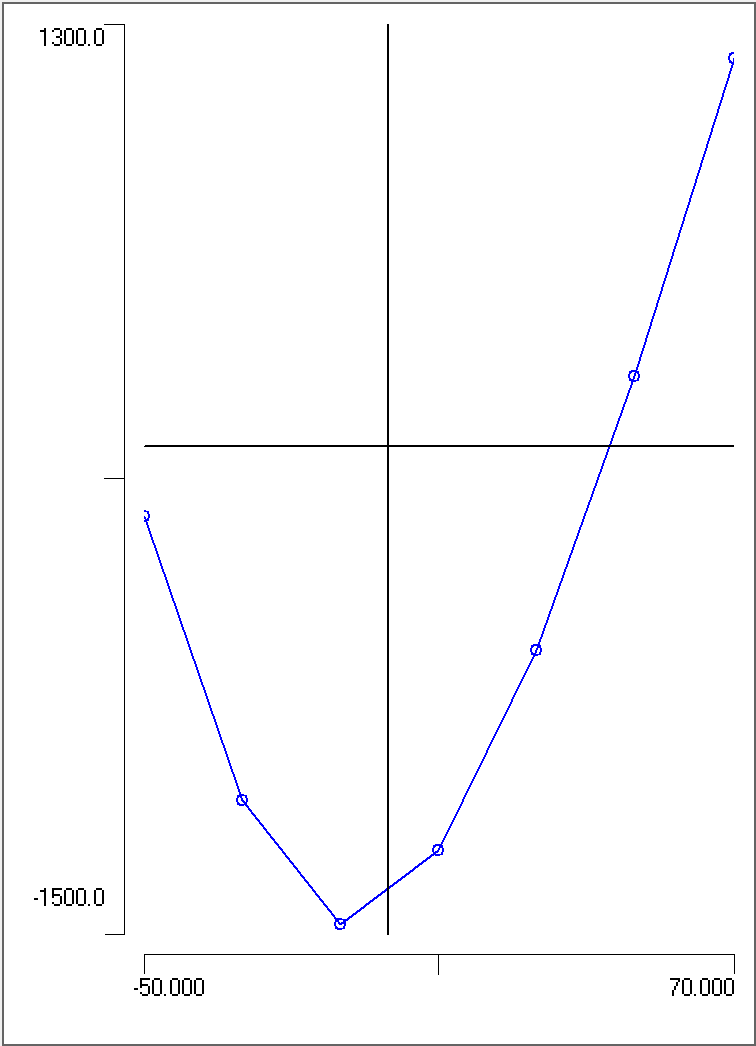
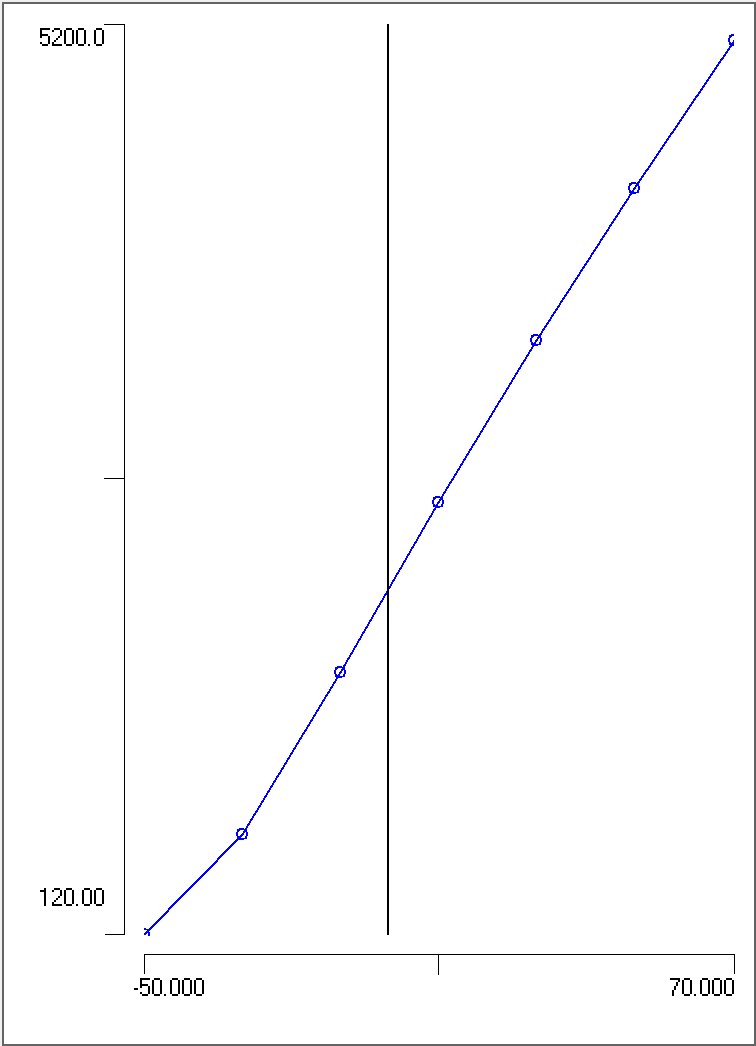
 ).
).